
Hace unos 4000 millones de años, cuando la tierra estaba recién formada, los primeros organismos aparecieron en la faz de nuestro planeta. Debe haber sido en algún cuerpo de agua – tal vez el mar, tal vez una charca. Con seguridad no lo sabemos, y las preguntas que tenemos son muchas. ¿Cómo eran los primeros organismos? ¿Dónde se desarrollaron? ¿Cómo se formaron? ¿Cómo vivían? Será imposible aclarar estas dudas de manera concluyente: el tiempo trascurrido desde aquel entonces borró prácticamente todas las trazas. Sin embargo, los investigadores han encontrado muchos indicios que, en su conjunto, nos dan una idea de cómo apareció la vida terrestre. En esta entrega veremos cuáles son los primeros organismos de los que tenemos constancia.
Para el estudio de las formas de vida del pasado, los paleontólogos – los científicos que se dedican a eso – suelen utilizar principalmente los restos fósiles de estos organismos. Estos restos consisten mayormente en lo que quedó de las partes duras del organismo: huesos, conchas, dientes… Las partes blandas de los organismos tienen una probabilidad extremamente baja de ser fosilizadas. Por tanto, hay un problema grave a la hora de estudiar los primeros organismos terrestres: al tratarse de organismos unicelulares sin partes duras, su potencial de ser fosilizados es mínimo, y es preciso de recurrir a otros métodos para su estudio. Veamos cuáles son estos métodos que nos permiten investigar los primeros organismos.

Estromatolitos
Aunque no haya partes duras de los primeros organismos que puedan haberse fosilizado, hay algo que dejaron algunos de ellos que sí podemos estudiar: los estromatolitos. Los estromatolitos (del griego στρώμα (strōma) = cama/alfombra y λίθος (lithos) = piedra) son estructuras minerales de forma globular, cónica o de hongo, con alturas que varían en escala de centímetros hasta decímetros, y que se encuentran en aguas someras (ver imagen). Su estructura interna es laminada; las láminas reflejan el crecimiento del estromatolito. Los estromatolitos modernos son el producto de la actividad de colonias de microbios (especialmente cianobacterias): la laminación es el resultado de la producción, captura y fijación de partículas carbonatadas por las biopelículas de las bacterias. El espesor de las láminas es en el orden de pocos milímetros, y su forma es variada: puede ser plana, hemisférica o columnar. Hoy día, los estromatolitos se encuentran sólo en algunas zonas específicas, tales como la Bahía Shark en Australia, donde no tienen depredadores por la alta salinidad del agua, pero en los inicios de la vida terrestre eran más comunes, y sus estructuras, conservadas en las rocas, son muy características (ver imágenes)[1]. De esta manera, la identificación de estromatolitos en rocas antiguas atestigua a la presencia de colonias de organismos bacterianos, aunque es preciso ser cautelosos y, antes de concluir que alguna estructura es un estromatolito, descartar que se haya formado por algún otro proceso[2].
Los estromatolitos actuales son el resultado de la presencia de colonias de bacterias de muchos tipos[3], y por analogía se considera que los estromatolitos antiguos lo son también. Sin embargo, no se ha podido confirmar con seguridad que es así, ya que no se ha encontrado bacterias fosilizadas en los estromatolitos – algo que es casi imposible, ya que, como vimos arriba, los organismos unicelulares tienen una probabilidad muy baja de ser fosilizados. Pero hay buenos indicios. Por ejemplo, los isótopos del azufre en el material orgánico que se encontró en unos estromatolitos de hace 3450 millones de años tienen una proporción igual a la que, hoy día, únicamente producen las bacterias que metabolizan azufre (en lugar de oxígeno); lo que indica la probable presencia de tales bacterias en los estromatolitos, y por ende un origen biogénico[4].
Los estromatolitos más viejos conocidos se encontraron en Groenlandia y datan de hace 3700 millones de años, o sea, de mediados del Arcaico, unos 800 millones de años después de la formación de la tierra[5] (ver imagen).


Con el paso del tiempo, los estromatolitos se volvieron más diversos. Hace 3430 millones de años, a mediados del Arcaico, ya formaron arrecifes (ver la imagen de la línea de tiempo, abajo). Su diversidad y abundancia eran más grandes en aquellos ambientes marinos costeros donde no había aportación ni de arcillas ni de arena, y donde no había fuentes calientes; y se reducían considerablemente en aguas más profundas[6]. Tapices de algas (hermanos de los estromatolitos, pero más planos) de hace 3480 millones de años parecen haberse formado en sabjas, o sea, ambientes costeros áridos y cálidos[7]. También se ha hallado estromatolitos de la misma edad asociados con depósitos de fuentes termales formados en un ambiente terrestre, lo que representaría la primera vida en tierra firme conocida[8].
El hecho de que los estromatolitos suelen ocurrir en aguas someras, sugiere que las bacterias que los formaron aprovechaban la luz del sol como fuente de energía: o sea, ya en el Arcaico puede haberse dado el proceso de fotosíntesis, la conversión de luz solar en energía[9]. Otra evidencia de actividad de fotosíntesis, ya en los primeros estromatolitos, se obtuvo analizando, en extensiones de estromatolitos cónicos fósiles, el espaciamiento entre los mismos, que resultó ser de algunos centímetros. En experimentos con estromatolitos actuales se encontró una relación entre su espaciamiento y la duración del ciclo de actividad de los microbios (un período de actividad más prolongado requiere de un mayor espaciamiento para que los microbios puedan obtener suficiente alimentación del agua). El espaciamiento observado en los estromatolitos antiguos indica un ciclo de actividad con una duración de alrededor de 20 horas, lo que es acorde con la duración de un día en la tierra joven (tal como vimos en la entrega anterior, la rotación de la tierra joven era más rápida que ahora). Al haber una relación entre el ciclo de actividad de los microbios de los estromatolitos y el ciclo de día y noche, es de suponer que los microbios aprovecharon la luz del sol para sus requerimientos energéticos, es decir que ya hubo fotosíntesis[10].
Desde mediados del Arcaico los estromatolitos se volvieron más y más comunes[11], de manera que se convirtieron en los íconos del Arcaico (ver la figura en el encabezado de esta entrega).

Microfósiles
Aunque los estromatolitos sean los restos más llamativos de los organismos del Arcaico y nos cuenten acerca de la primera vida en la tierra, mejor sería encontrar fósiles de los organismos mismos. Por tratarse de organismos unicelulares, micrométricos y muy antiguos, no es fácil encontrar tales fósiles y poder estar seguros de que sean de verdad restos de organismos y no – por ejemplo – algún crecimiento de minerales (esto ocurrió en el caso de lo que se creyó que eran microfósiles de hace 3500 millones de años[12], pero que parecen ser filamentos minerales[13], aunque el debate sigue abierto[14]).
Pero se ha encontrado ejemplares que tienen toda la pinta de representar microfósiles de las primeras bacterias. Hasta hace pocos años no se conocían microfósiles inequívocos del Arcaico. Sólo en rocas del eón sucesivo, el Proterozoico, se encontraron microfósiles aceptados como tal por la mayoría de investigadores. Entre estos se encuentran los microfósiles hallados en el sílex de Gunflint (Canadá), con una edad de 1900 millones de años[15].
Sin embargo, hace unos años se publicaron hallazgos de lo que parecen ser microfósiles verídicos del Arcaico. De finales del Arcaico datan estructuras tubulares micrométricas, encontradas en sedimentos lacustres en Sudáfrica, que tienen un gran parecido con algas conocidas del Proterozoico[16]. Aun más viejos son los restos orgánicos globulares y tubulares, micrométricos, encontrados en rocas sedimentarias formadas en antiguas playas de hace 3400 millones de años, en Australia: tienen la forma de células, y además los valores isotópicos del carbono y azufre encontrado en estos restos son valores típicos de los productos de procesos biológicos (ver figura). Por tanto, luce probable que se trata de las células de organismos unicelulares que obtenían su energía reduciendo azufre[17] (en aquel tiempo aun no había oxígeno en la atmósfera).

Aun más viejos serían, de ser confirmadas como microorganismos, las estructuras tubulares de hematita (óxido de hierro), con una edad de entre 3770 y 4280 millones de años, que corresponde al inicio del Arcaico o, inclusive, finales del eón anterior, el Hádico (ver la imagen de la línea de tiempo). Estas estructuras se encontraron en Canadá, en rocas formadas en un ambiente marino profundo, probablemente relacionado a fuentes hidrotermales[18]. Tal como veremos en la próxima entrega de este blog, hay una hipótesis que dice que la vida terrestre se originó en la cercanía de fuentes hidrotermales, o sea, de agua caliente rica en minerales. De confirmarse estas estructuras tubulares como microfósiles, serían los más viejos encontrados hasta la fecha, y darían testimonio de una aparición de la vida cuando la tierra todavía era joven y las condiciones poco propicias: entre otras cosas, nuestro planeta estaba todavía siendo bombardeada por meteoritos (ver la entrega anterior de este blog).
Isótopos
Ya vimos cómo, para determinar si un posible resto de organismos de verdad es orgánico, los investigadores a menudo se apoyan en la información de los isótopos, ya que pueden ayudar a diferenciar entre moléculas formadas por procesos biológicos y aquellas que se formaron de otra manera.
Muchos elementos tienen múltiples isótopos, o sea variantes del mismo elemento pero con masas ligeramente distintas, debido a diferentes cantidades de neutrones en el núcleo; algunos isótopos son estables, otros radioactivos, o sea, se desintegran a lo largo del tiempo. En la entrega anterior ya se habló de los isótopos radioactivos, que nos permiten datar las rocas, y de los isótopos estables de algunos elementos, tales como el azufre, que tienen relevancia para entender el entorno químico en la superficie terrestre. Pero los isótopos estables de ciertos elementos también son de gran utilidad para investigar la presencia de la vida.
El principal elemento cuyos isótopos tienen relevancia para la vida, es el carbono. Todos los compuestos orgánicos, de los que se componen los organismos terrestres, contienen carbono. En la ausencia de microfósiles confiables en las rocas más antiguas del planeta, se ha propuesto utilizar la presencia de material carbonoso con una baja relación isotópica del carbono como indicación de actividad biológica. El elemento carbono ocurre en la naturaleza con tres isótopos, que se distinguen por la cantidad de neutrones en el núcleo. Dos de los isótopos son estables: carbono-12 (12C [19]) y carbono-13 (13C); el primero es el más abundante[20]. Los procesos biológicos evitan el uso del 13C para la elaboración de los compuestos orgánicos, lo que resulta en una relación entre 12C y 13C muy alta en estos compuestos, los cuales por tanto se llaman isotópicamente ligeros[21]. Puesto que no conocemos otros procesos que resultan en carbono isotópicamente tan ligero, se está utilizando la ocurrencia de carbono isotópicamente ligero en las rocas antiguas como evidencia, aunque fuera indirecta, de la presencia de procesos biológicos y por ende, alguna forma de vida. Esto es lo que ha conducido a algunos investigadores proponer la presencia de vida en nuestro planeta antes de hace 3800 millones de años, basándose en inclusiones de material carbonoso muy ligero isotópicamente, en rocas de esa edad encontradas en Groenlandia[22]. Inclusive, la presencia de grafito isotópicamente ligero en inclusiones dentro de cristales de zircón de Australia con una edad de 4100 millones de años (finales del Hádico), pudiera indicar la presencia de organismos – lo que sería la vida más antigua conocida[23]. Sin embargo, este razonamiento no necesariamente es válido: muchas de las rocas antiguas se formaron por procesos volcánicos, en ambientes donde es imposible que haya habido vida; en tales rocas, el carbono isotópicamente ligero difícilmente puede tener un origen biológico[24]. Lo mismo ocurre con el grafito (un mineral carbonoso) que se ha encontrado en rocas antiguas en Groenlandia: parece ser isotópicamente ligero, pero se formó por procesos que ocurren a gran profundidad en la corteza terrestre, y por tanto no tiene relación alguna con procesos biológicos[25].
Especialmente cuando se utilizan en combinación con otros métodos que pudieran indicar que restos carbonosos son de verdad de origen biológico (por ejemplo, análisis químicos o microscópicos), los isótopos de carbono pueden ser de gran utilidad. Por ejemplo, filamentos carbonosos de hace 3500 millones de años, encontrados en depósitos formados alrededor de fuentes hidrotermales sulfurosas en Australia, tienen – entre otros indicios que apuntan a un origen biológico – valores isotópicos muy bajos y por tanto pueden ser considerados como restos microbianos[26]. Lo mismo ocurre con posibles restos de organismos unicelulares con forma de huso de hace 3000 millones de años, cuyos isótopos de carbono indican un origen biológico; esto sugiere que se trata de organismos planctónicos[27]. Asimismo, la combinación de isótopos ligeros con otros indicios de vida en múltiples afloramientos de rocas de origen marino con unas edades de entre 3500 y 3200 millones de años da mayor peso a la aseveración que hubo organismos vivos en aquellos tiempos[28].
Otros elementos cuyos isótopos estables son de utilidad para identificar los procesos biológicos, incluyen el azufre, el nitrógeno e inclusive algunos metales. En el caso de los isótopos estables del azufre, los cambios en la proporción de estos isótopos (lo que se denomina el fraccionamiento) se deben a factores ambientales y al metabolismo del organismo. Variaciones temporales en el fraccionamiento del azufre en los restos de organismos antiguos pueden, por ejemplo, revelar la alternancia de períodos de abundancia de alimentos y períodos de escasez[29].
El nitrógeno es un elemento clave para los organismos, al ser un componente de los aminoácidos y las proteínas. Se encuentra en grandes cantidades en la atmósfera, pero en una forma que los organismos no pueden usar. Para que el nitrógeno sea utilizable, tiene que ser fijado: al hidrógeno, formándose amonio, o al oxígeno, formándose nitrato. Son pocos los organismos capaces de fijar el nitrógeno: sólo algunos grupos de bacterias y ciertas arqueobacterias[30]. El elemento nitrógeno tiene dos isótopos estables: 14N y 15N. Así como en el caso de los isótopos de carbono, los organismos prefieren utilizar el isótopo más ligero, en este caso 14N, de manera que por lo general los organismos tienen nitrógeno isotópicamente ligero, aunque haya muchos variables que influyen sobre esta relación entre 14N y 15N [31]. En rocas del Arcaico tardío, arcillas con alto contenido de material orgánico de origen bacteriano, variaciones en la relación entre los isótopos de nitrógeno indican un período de formación de nitratos (o sea, de fijación del nitrógeno en la presencia de oxígeno), lo que implica una fase de disponibilidad de oxígeno en el ambiente[32], antes de la gran oxidación de la cual hablaremos en una entrega venidera.
Otro elemento clave para los organismos es el fósforo. Desafortunadamente, el fósforo sólo tiene un isótopo estable, así que no se puede estudiar de la misma manera que los isótopos de carbono, nitrógeno etc. Sí se pudo determinar que, hasta hace unos 800 millones de años, la cantidad de fósforo en los océanos fue relativamente baja, lo que puede haber limitado la biomasa, o sea, la cantidad total de organismos[33].
Entre los metales son de interés el hierro y, especialmente, el níquel. El níquel es un elemento que, aunque fuera en cantidades mínimas, es imprescindible para los organismos metanogénicos, que obtienen su energía convirtiendo dióxido de carbono e hidrógeno en metano[34]. Estos organismos, de los que volveremos a hablar abajo, eran la forma de vida dominante antes de que apareciera la fotosíntesis a finales del Arcaico. En el material orgánico encontrado en las rocas antiguas, un enriquecimiento en níquel isotópicamente ligero puede indicar la presencia de tales organismos metanogénicos, y así ayudar a entender cuándo y cómo éstos se desarrollaron[35].
Biomarcadores
A falta de restos de células de organismos enteros, pueden servir como evidencia de la presencia de organismos los restos de moléculas biológicas, que en su momento formaban parte de las células. Estos restos de moléculas biológicas se llaman biomarcadores.
Biomarcadores son compuestos orgánicos que indican la presencia de algún proceso biológico o, sea, la presencia de organismos[36]. Al hallar en rocas antiguas algún biomarcador, podemos concluir que existieron organismos vivos en la época que se formaron estas rocas. Los organismos mismos probablemente no dejaron restos fósiles, pero sí pudieron haber dejado rastros de su existencia en la forma de moléculas específicas de los organismos vivos. Inclusive, a veces es posible relacionar un biomarcador a algún proceso específico o a algún grupo de organismos en particular.
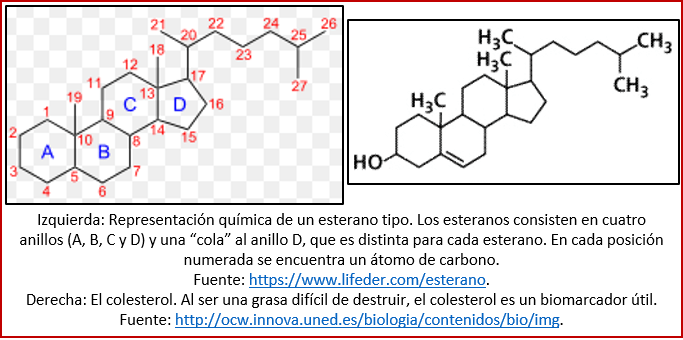
Un ejemplo de los biomarcadores son los esteranos (ver figura). Son moléculas orgánicas formadas como resultado de la degradación de esteroides. Estos compuestos son utilizados principalmente en las células eucariontes (o sea, las que tienen un núcleo). Unos esteroides conocidos son el colesterol, la cortisona y la testosterona. Así que, si se encuentra un esterano en rocas antiguas, se sabe no sólo que hubo organismos cuando se depositó esta roca, sino también que estos organismos eran probablemente eucariontes. Puesto que, tal como veremos abajo, los primeros organismos fueron procariontes (organismos unicelulares sin núcleo), los biomarcadores dejados por éstos fueron otros, y la aparición de los esteranos sirve para datar los primeros eucariontes [37].
Entre los biomarcadores que se han encontrados en las rocas del Arcaico, figuran ciertos esteranos que aparecen desde el final del Arcaico, y que pudieran estar relacionados al proceso de fotosíntesis, o sea, con la producción de oxígeno. También se encontraron carotenoides de la misma edad, compuestos también relacionados a la fotosíntesis, en ciertas bacterias[38]. Por su relevancia para datar los primeros organismos capaces de realizar fotosíntesis[39], volveremos a hablar de estos biomarcadores en una entrega venidera, dedicada a la aparición de la fotosíntesis.
Los biomarcadores son un componente del petróleo. En rocas donde hay muchos biomarcadores, es posible que ocurrió la generación o la acumulación de petróleo (que se forma a partir de acumulaciones de restos de organismos principalmente unicelulares). De hecho, en regiones como Australia occidental o Sudáfrica, se ha encontrado en rocas del Arcaico no sólo biomarcadores, sino también restos de petróleo, en la forma de gotas o bitumen. La ocurrencia más antigua de petróleo es de gotas encontradas en Australia, en rocas con una edad de más de 3460 millones de años. Se piensa que estas gotas se pudieron formar en esa misma roca, lo que sería otra evidencia para una abundancia importante de bacterias en los océanos del Arcaico medio[40].
La naturaleza de los primeros organismos
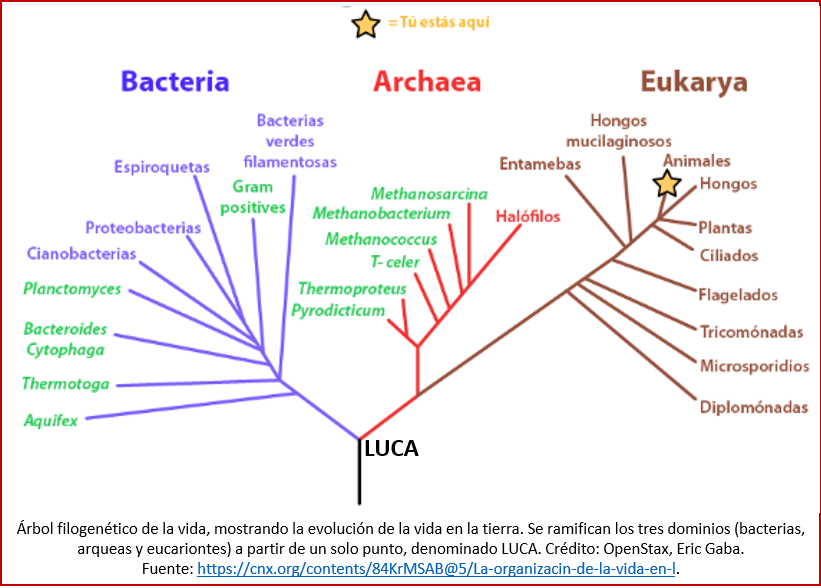
¿Cuáles fueron estos primeros organismos? Todos los organismos que conocemos caen en uno de los tres dominios de la vida que han sido identificados[41] (ver figura): los eucariontes, las arqueas, y las bacterias (los últimos dos conforman los procariontes)[42]. Los eucariontes incluyen todos aquellos organismos cuyas células tienen un núcleo. Las bacterias son organismos unicelulares sin núcleo definido (por eso pertenecen a los procariontes). Las arqueas se parecen a primera vista a las bacterias, pero son fundamentalmente distintas: las arqueas poseen genes y varias rutas metabólicas que son más cercanas a las de los eucariontes, y en otros aspectos son únicas, tales como la composición de sus membranas celulares. Las arqueas explotan una variedad de recursos mucho mayor que los eucariontes, desde compuestos orgánicos comunes como los azúcares, hasta el uso de amoníaco, iones de metales o incluso hidrógeno como nutrientes. Las arqueas existen en una gran variedad de hábitats; muchas arqueas son extremófilas, y este tipo de hábitat fue visto históricamente como su nicho ecológico. De hecho, algunas arqueas sobreviven a altas temperaturas, como la cepa 121 de Geogemma barossii, a menudo por encima de 100 °C, como las que hay en los géiseres, chimeneas mineralizadas, y pozos de petróleo[43].
Por tanto, se considera que los primeros organismos, que vivían mucho antes de la aparición del oxígeno en la atmósfera terrestre, pueden haber pertenecido al dominio de las arqueas, y que éstos eran metanogénicos, o sea, tenían un metabolismo basado en la producción de metano a partir de hidrógeno y CO2 [44] (hoy día todavía existen arqueas metanogénicas en ambientes sin oxígeno[45]). Inicialmente se pensó que los organismos más viejos eran cianobacterias, que son los únicos procariontes capaces de fotosíntesis[46], pero ahora se sabe que las arqueas son más primitivas. Entre las arqueas, las extremófilas parecen estar entre las más antiguas[47], de manera que se postuló que la vida terrestre se originó en ambientes de fuentes calientes (de que hablaremos la próxima entrega), a temperaturas altas o tal vez más moderadas[48].
La evidencia más antigua de metanogénesis data de hace 3460 millones de años, del Arcaico temprano. Se trata de metano entrampado en minúsculas burbujas dentro de rocas de esa edad; este metano fue analizado en el laboratorio y resultó ser extremadamente ligero isotópicamente – lo que implica que se formó debido a un proceso biológico. Esto indica no sólo que ya hubo vida en la tierra en aquel tiempo, sino también que hubo organismos metanogénicos[49].
Estudios recientes indican que las primeras arqueas, de hace más de 3500 millones de años, eran en efecto anaeróbicas, o sea, que vivían en un ambiente sin oxígeno, que tenían un genoma pequeño (posteriormente ampliado por incorporaciones de material genético de, especialmente, bacterias[50]), y que probablemente obtenían su energía convirtiendo CO2 en acetato[51].
Otros organismos posiblemente muy viejos son los virus. Tal como vimos en una anterior entrega de este blog (¿Qué es la vida?, del 15 mayo 2018), los virus dependen de otros organismos por su replicación y por tanto se supone que aparecieron después de los primeros organismos. Pero es posible que se desarrollaron muy temprano, ya en los tiempos de las primeras arqueas[52].
Reconstrucción de moléculas biológicas
El estudio de las moléculas en organismos procariontes actuales ha permitido reconstruir algunas de las moléculas y funciones que deben haber estado presentes en los primeros organismos: al comparar moléculas parecidas en organismos distintos, ha sido posible elaborar, en algunos casos, un árbol genealógico de estas moléculas en el tiempo, y determinar cómo pueden haber sido estas moléculas en los primeros organismos. Esto nos permite obtener una idea de cómo eran estas formas de vida ancestral, y entender mejor el desarrollo de la vida en nuestro planeta y su evolución.
Entre los compuestos estudiados se encuentran las proteínas. Las proteínas, aparte de tener cada una su composición química específica, tienen también sus estructuras características, ya que cada proteína se dobla de una manera propia. Las proteínas pueden ser agrupadas por manera de doblarse. Estas maneras de doblarse resultan ser muy estables en el tiempo: la composición de una proteína puede cambiar a lo largo del tiempo, pero su manera de doblarse es prácticamente inmutable. Esto permite reconstruir el árbol evolucionario de las proteínas, y relacionarlo a la evolución de la vida[53].
Las proteínas, así como el material genético de todos los organismos terrestres, están compuestos de aminoácidos. Algunos aminoácidos tienen una composición química simple (tan simple que inclusive se han encontrado en el espacio, lejos de cualquier forma de vida; ver la entrega del 26 agosto 2017, “El preludio de la vida (6): Semillas universales”, de este blog), y se considera que los primeros organismos utilizaron estos aminoácidos sencillos para su construcción. Con el pasar del tiempo, los organismos deben haberse vuelto más sofisticados y capaces de construir aminoácidos más complejos. Uno de estos, que sólo se logra fabricar con la ayuda de ciertas proteínas, es el triptófano. El triptófano es uno de los aminoácidos menos comunes, y parece que fue el último en aparecer; esto se sabe porque se ha logrado reconstruir varias proteínas primordiales, que contienen todos los aminoácidos menos el triptófano[54].
La proteína necesaria para la fabricación del triptófano, llamada triptófano sintasa, ya era una proteína compleja cuando apareció por primera vez, parecida en complejidad a la proteína que hoy día es responsable de la producción del triptófano. En otras palabras, los organismos primordiales se volvieron complejos bastante rápidamente; si en sus inicios eran primitivos y simples, pronto se volvieron complejos[55].
El dominio de los procariontes
De la vida antes de LUCA (el ancestro común de todos los organismos actuales – ver abajo) sabemos muy poco. Aparecieron las grandes innovaciones que hicieron posible la vida que conocemos hoy día: específicamente, el “triunvirato biológico” del envoltorio celular (y el mecanismo de división celular), el metabolismo, y el material genético[56].
Las condiciones de vida no fueron fáciles para los primeros procariontes. Una radiación ultravioleta intensa y temperaturas más o menos elevadas deben haber causado daños en el material genético de los organismos, lo que por un lado puede haber resultado en una tasa de mutaciones alta y por ende un ritmo de evolución elevado[57], pero por otro lado puede haber causado daños en el material genético, que tuvieron que ser contrarrestados por mecanismos reparadores[58]. Además, las temperaturas elevadas causan que se sueltan un poco los filamentos del ADN y el ARN en el genoma, de manera que se pueden insertar con mayor facilidad trozos de material genético denominados intrones; estos suelen facilitar el corte del ADN y ARN, haciendo posible los cambios genéticos[59].
La ausencia de oxígeno durante el Hádico y el Arcaico implica que los primeros organismos tuvieron que obtener su energía de otras fuentes. Arriba ya se mencionaron las arqueas metanogénicas, que obtienen su energía al convertir CO2 y acetato en metano (y agua) [60]. Otra fuente de energía es la oxidación de sulfuro, o sea la conversión de azufre reducido, por ejemplo sulfuro de hidrógeno (el gas que da a los huevos podridos su olor típico), en sulfatos. Probables fósiles de bacterias de hace 2520 millones de años, o sea del final de Arcaico, contienen una variedad de isótopos del azufre tal que se supone que estas bacterias procesaban sulfuros[61].
Otro aspecto importante de la vida es la capacidad de fijar CO2 en la forma de compuestos orgánicos necesarios para el desarrollo y mantenimiento del organismo. En la actualidad, cada organismo tiene un mecanismo para hacer esto; en total se conocen seis mecanismos distintos. En los organismos primordiales, sin embargo, existieron sólo algunos mecanismos y en cada organismo había múltiples mecanismos actuando: una redundancia, probablemente causada por una confiabilidad escasa de cada uno de los mecanismos. Sólo más tarde, cuando se ramificaron los distintos dominios de la vida de los organismos ancestrales, los mecanismos de fijación del CO2 se diversificaron y mejoraron[62].
LUCA
Los tres dominios de la vida terrestre (ya los vimos arriba: las bacterias, las arqueas y los eucariontes) tienen en común ciertas características, que hacen pensar que deben descender de algún ancestro común. Esta forma ancestral hipotética se denomina el LUCA: Last Universal Common Ancestor (último antepasado común universal). No conocemos ningún resto fósil del LUCA, pero podemos deducir varias cosas acerca de este gracias a la reconstrucción de las moléculas biológicas a partir de los procariontes (y eucariontes) actuales[63].
En primer lugar, todos los organismos actuales tienen un genoma basado en el ADN. Por tanto, es de suponer que el LUCA lo haya tenido también. Utilizando este mismo razonamiento, se ha elaborado listas de genes que son iguales o parecidos en los tres dominios de la vida, y se ha sugerido que ya existían en el LUCA. La aplicación de esta técnica, sin embargo, se complica porque ha habido mucho intercambio de material genético entre especies: las bacterias suelen intercambiar material genético, lo que complica la construcción de árboles genealógicos de las especies, y por ende la determinación de cómo lucía el LUCA[64]. Afortunadamente, gracias a la grande base de datos genéticos de organismos de todos los dominios de la vida, así como la actual capacidad de computación, se ha podido reconstruir las proteínas primordiales y determinar cuáles pueden haber sido presentes en el LUCA. Con base en las funciones de estas proteínas, se concluyó que el LUCA vivió antes de que la atmósfera tuviera oxígeno. Sin embargo, no sabemos cuáles fueron los genes que tuvo el LUCA y que no fueron transmitidos a todos los dominios de la vida, de manera que la información que tenemos acerca del LUCA es incompleta[65].
El LUCA probablemente era un organismo metanogénico, que obtenía su energía a partir de hidrógeno. Algunas de las proteínas identificadas pueden haber estado involucradas en la adaptación a temperaturas altas, lo que sugiere que el LUCA estaba adaptado a un ambiente hidrotermal, tal vez fuentes submarinas donde brotan aguas calientes cargadas de minerales[66]. Sin embargo, un análisis del ARN ribosomal (que puede haber servido como base del genoma, antes de que apareciera el ADN (de esto hablaremos en la próxima entrega) sugiere que la temperatura del agua no fue tan extrema: tal vez no más de 50°C [67].
Además, se considera que la membrana celular del LUCA, al ser el ancestro directo de dos dominios, las arqueas y las bacterias (las bacterias fueron las primeras a separarse de las arqueas, mientras que los eucariontes hicieron lo mismo más tarde[68]), puede haber tenido características tanto de las bacterias como de las arqueas. Tal como ya se indicó arriba, las membranas de estos dos dominios de la vida son distintas. Se ha determinado que procariontes con membranas mixtas son estables, y por tanto es posible que el LUCA haya tenido una membrana híbrida, que posteriormente dio origen a las membranas diferentes de las arqueas y las bacterias[69].
No sabemos cuándo vivió el LUCA. Obviamente debe haber sido después de la aparición de los primeros organismos en la tierra, y antes de que se separaran las arqueas y bacterias en dos dominios. Se estima que puede haber sido hace 3500-3800 millones de años[70]. En la línea de tiempo (ver imagen arriba) se coloca LUCA tentativamente en 3500 millones de años.
Tampoco sabemos si el LUCA fue de verdad un solo individuo. Ciertos investigadores prefieren considerar al LUCA como un grupo de individuos, que en su conjunto dieron origen a todas las especies posteriores[71].
Conclusión
En esta entrega vimos cuáles son los primeros organismos de los que tenemos constancia; pero también vimos cuán difícil es determinar exactamente cuándo apareció la vida, y cómo la poca evidencia hace difícil determinar con seguridad cuáles fueron los primeros organismos. Las líneas de evidencia se basan a menudo en indicios indirectos, tales como biomarcadores e isótopos. Microfósiles son escasos, y su interpretación cómo organismos es a menudo ambigua. Lo que sí nos da cierto apoyo, son las estructuras tales como los estromatolitos, que se pueden considerar el resultado de la actividad de colonias de organismos.
En algún momento, tal vez hace 3500-3800 millones de años, empezaron a diferenciarse distintos tipos de organismos, que dieron origen a los tres dominios de la vida. Antes de este momento pueden haberse dado ramificaciones también, pero estas se extinguieron más o menos pronto y no tenemos evidencias de su existencia. El hipotético organismo a partir del cual empezó la ramificación de los tres dominios, ha sido denominado el LUCA.
Después del LUCA empezaron a diferenciarse los dominios de la vida: primero se separaron las bacterias y las arqueas, mientras que los eucariontes se separaron más tarde de las arqueas. Los eucariontes dieron origen, después de un período largo de poca evolución aparente, a los organismos multicelulares. Los procariontes se han ido diversificando de manera continua en el tiempo, formando una multitud de linajes que han perdurado hasta el día de hoy[72]. De esta historia volveremos a hablar en unas entregas futuras.
Esto es, en pocas palabras, lo que sabemos de la aparición de los primeros organismos. Pero poco nos dice esto acerca de cuáles fueron los procesos que causaron la transformación de materia muerta en los organismos vivos. ¿Qué sabemos de esto? En la próxima entrega veremos cómo pudiera haber aparecido esta primera vida.
Nota: la imagen en el encabezado de esta entrega es una impresión artística de cómo puede haber lucido la superficie terrestre a finales del Arcaico. Estromatolitos crecían en las orillas de los mares. Crédito: Peter Sawyer / Smithsonian Institution. Fuente: www.astrobio.net/also-in-news/life-earths-primordial-sea-starved-sulfate.
[1] Ver: https://es.wikipedia.org/wiki/Estromatolito.
[2] Ver: www.youtube.com/watch?v=HIniR9KOA1E.
[3] Klatt, C.G. y otros, 2011. Community ecology of hot spring cyanobacterial mats: predominant populations and their functional potential. The ISME Journal, 5, 1262-1278. https://www.nature.com/articles/ismej201173.pdf. Melendrez, M.C., Lange, R.K., Cohan, F.M. y Ward, D.M., 2011. Influence of molecular resolution on sequence-based discovery of ecological diversity among Synechococcus populations in an alkaline siliceous hot spring microbial mat. Applied and Environmental Microbiology, 77 (4), p. 1359-1367. http://aem.asm.org/content/77/4/1359.full.pdf.
[4] Bontognali, T.R.R., Sessions, A.L., Allwood, A.C., Fischer, W.W., Grotzinger, J.P., Summons, R.E. y Eiler, J.M., 2012. Sulfur isotopes of organic matter preserved in 3.45-billion-year-old stromatolites reveal microbial metabolism. Proceedings of the National Academy of Sciences, 109 (38) 15146-15151. www.pnas.org/content/109/38/15146.
[5] Nutman, A.P., Bennett, V.C., Friend, C.R.L., Van Kranendonk, M.J. y Chivas, A.R., 2016. Rapid emergence of life shown by discovery of 3,700-million-year-old microbial structures. Nature, 537, 535-538. www.nature.com/articles/nature19355.
[6] Allwood, A.C., Walter, M.R., Kamber, B.S., Marshall, C.P. y Burch, I.W., 2006. Stromatolite reef from the Early Archaean era of Australia. Nature, 441, 714-718. www.nature.com/articles/nature04764. Allwood, A.C., Walter, M.R., Burch, I.W. y Kamber, B.S., 2007. 3.43 billion-year-old stromatolite reef from the Pilbara Craton of Western Australia: Ecosystem-scale insights to early life on Earth. Precambrian Research, 158 (3-4), 198-227. www.sciencedirect.com/science/article/pii/S0301926807001234. Allwood, A.C., Grotzinger, J.P., Knoll, A.H., Burch, I.W., Anderson, M.S., Coleman, M.L. y Kanik, I., 2009. Controls on development and diversity of Early Archean stromatolites. Proceedings of the National Academy of Sciences, 106 (24), 9548-9555. www.pnas.org/content/pnas/106/24/9548.full.pdf.
[7] Noffke, N., Christian, D., Wacey, D. y Hazen, R.M., 2013. Microbially induced sedimentary structures recording an ancient ecosystem in the ca. 3.48 billion-year-old Dresser Formation, Pilbara, Western Australia. Astrobiology, 13 (12), 1103-24. www.liebertpub.com/doi/pdf/10.1089/ast.2013.1030.
[8] Djokic, T., Van Kranendonk, M.J., Campbell, K.A., Walter, M.R. y Ward, C.R., 2017. Earliest signs of life on land preserved in ca. 3.5 Ga hot spring deposits. Nature Communications, 8, 15263. www.nature.com/articles/ncomms15263.
[9] Tice, M.M. y Lowe, D.R., 2004. Photosynthetic microbial mats in the 3,416-Myr-old ocean. Nature, 431, 549-552. www.nature.com/articles/nature02888.
[10] Petroff, A.P., Sim, M.S., Maslov, A., Krupenin, M., Rothman, D.H. y Bosak, T., 2010. Biophysical basis for the geometry of conical stromatolites. Proceedings of the National Academy of Sciences, 107 (22), 9956-9961. www.pnas.org/cgi/doi/10.1073/pnas.1001973107.
[11] Schopf, J.W., 2006. Fossil evidence of Archaean life. Philosophical Transactions of the Royal Society B, 361, 869-885. http://rstb.royalsocietypublishing.org/content/361/1470/869. Schopf, J.W., Kudryavtsev, A.B., Czaja, A.D. y Tripathi, A.B., 2007. Evidence of Archean life: Stromatolites and microfossils. Precambrian Research, 158, 141-155. www.sciencedirect.com/science/article/pii/S0301926807001209.
[12] Schopf, J.W. y Packer, B.M., 1987. Early Archean (3.3-billion to 3.5-billion-year-old) microfossils from Warrawoona Group, Australia. Science, 237 (4810), 70-73. http://science.sciencemag.org/content/237/4810/70. Schopf, J.W., Kudryavtsev, A.B., Agresti, D.G., Wdowiak, T.J. y Czaja, A.D., 2002. Laser–Raman imagery of Earth’s earliest fossils. Nature, 416, 73-76. www.nature.com/articles/416073a. Schopf, J.W., 2004. Geochemical and submicron-scale morphologic analyses of individual Precambrian microorganisms. The Geochemical Society Special Publications, 9, 365-375. www.sciencedirect.com/science/article/pii/S1873988104800269.
[13] Brasier, M.D., Green, O.R., Jephcoat, A.P., Kleppe, A.K., Van Kranendonk, M.J., Lindsay, J.F., Steele, A. y Grassineau, N.V., 2002. Questioning the evidence for Earth’s oldest fossils. Nature, 416, 76-81. www.nature.com/articles/416076a. Marshall, C.P., Emry, J.R. y Olcott Marshall, A., 2011. Haematite pseudomicrofossils present in the 3.5-billion-year-old Apex Chert. Nature Geoscience, 4, 240–243. www.nature.com/articles/ngeo1084. Brasier, M.D., Antcliffe, J., Saunders, M. y Wacey, D., 2015. Changing the picture of Earth’s earliest fossils (3.5–1.9 Ga) with new approaches and new discoveries. Proceedings of the National Academy of Sciences, 112 (16), 4859-4864. www.pnas.org/content/112/16/4859.
[14] Schopf, J.W., Kitajima, K., Spicuzza, M.J., Kudryavtsev, A.B. Valley, J.W., 2018. SIMS analyses of the oldest known assemblage of microfossils document their taxon-correlated carbon isotope compositions. Proceedings of the National Academy of Sciences, 115 (1) 53-58. www.pnas.org/content/115/1/53.
[15] Wacey, D., McLoughlin, N., Kilburn, M.R., Saunders, M., Cliff, J.B., Kong, C., Barley, M.E. y Brasier, M.D., 2013. Nanoscale analysis of pyritized microfossils reveals differential heterotrophic consumption in the ∼1.9-Ga Gunflint chert. Proceedings of the National Academy of Sciences, 110 (20), 8020-8024. www.pnas.org/content/110/20/8020.
[16] Kaźmierczak, J., Kremer, B., Altermann, W. y Franchi, I., 2016. Tubular microfossils from ∼2.8 to 2.7 Ga-old lacustrine deposits of South Africa: A sign for early origin of eukaryotes? Precambrian Research, 286, 180-194. www.sciencedirect.com/science/article/pii/S0301926816301401.
[17] Wacey, D., Kilburn, M.R., Saunders, M., Cliff, J. y Brasier, M.D., 2011. Microfossils of sulphur-metabolizing cells in 3.4-billion-year-old rocks of Western Australia. Nature Geoscience, 4, 698-702. www.nature.com/articles/ngeo1238. Brasier, M.D.,, McLoughlin, N., Green, O. y Wacey, D., 2006. A fresh look at the fossil evidence for early Archaean cellular life. Philosophical Transactions of the Royal Society B, 361, 887-902. http://rstb.royalsocietypublishing.org/content/361/1470/887.
[18] Dodd, M.S., Papineau, D., Grenne, T., Slack, J.F., Rittner, M., Pirajno, F., O’Neil, J. y Little, C.T.S., 2017. Evidence for early life in Earth’s oldest hydrothermal vent precipitates. Nature, 543, 60-64. www.nature.com/articles/nature21377.
[19] El número (12 en este caso) se refiere a la suma de protones y neutrones en el núcleo del átomo.
[20] Ver: https://es.wikipedia.org/wiki/Is%C3%B3topo.
[21] Ver: https://en.wikipedia.org/wiki/%CE%9413C.
[22] Mojzsis, S.J., Arrhenius, G., McKeegan, K.D., Harrison, T.M., Nutman, A.P. y & Friend, C.R.L., 1996. Evidence for life on Earth before 3,800 million years ago. Nature, 384 (6604), 55-59. www.nature.com/articles/384055a0.
[23] Bell, E.A., Boehnke, P., Harrison, T.M. y Mao, W.L., 2015. Potentially biogenic carbon preserved in a 4.1 billion-year-old zircon. Proceedings of the National Academy of Sciences, 112 (47), 14518-14521. http://www.pnas.org/content/112/47/14518.
[24] Fedo, C.M., Whitehouse, M.J. y Kamber, B.S., 2006. Geological constraints on detecting the earliest life on Earth: a perspective from the Early Archaean (older than 3.7 Gyr) of southwest Greenland. Philosophical Transactions Royal Society B, 361, 851-867. http://rstb.royalsocietypublishing.org/content/361/1470/851.
[25] Van Zuilen, M.A., Lepland, A. y Arrhenius, G., 2002. Reassessing the evidence for the earliest traces of life. Nature, 384 (6604), 55-59. www.nature.com/articles/384055a0.
[26] Glikson, M., Duck, L.J., Golding, S.D., Hofmann, A., Bolhar, R., Webb, R., Baiano, J.C.F. y Sly, L.I., 2008. Microbial remains in some earliest Earth rocks: Comparison with a potential modern analogue. Precambrian Research, 164 (3-4), 187-200. www.sciencedirect.com/science/article/pii/S0301926808001083. Duck, L.J., Glikson, M., Golding, S.D. y Webb, R.E., 2007. Microbial remains and other carbonaceous forms from the 3.24 Ga Sulphur Springs black smoker deposit, Western Australia. Precambrian Research, 154 (3-4), 205-220. www.sciencedirect.com/science/article/pii/S0301926807000253.
[27] House, C.H., Oehler, D.Z., Sugitani, K. y Mimura, K., 2013. Carbon isotopic analyses of ca. 3.0 Ga microstructures imply planktonic autotrophs inhabited Earth’s early oceans. Geology, 41 (6), 651-654. https://pubs.geoscienceworld.org/gsa/geology/article-abstract/41/6/651/131229.
[28] Schopf, 2006. Ver nota 11.
[29] Wing, B.A. y Halevy, I., 2014. Intracellular metabolite levels shape sulfur isotope fractionation during microbial sulfate respiration. Proceedings of the National Academy of Sciences, 111 (51), 18116-18125. www.pnas.org/content/111/51/18116.
[30] Ver: https://es.wikipedia.org/wiki/Fijaci%C3%B3n_de_nitr%C3%B3geno; https://es.wikipedia.org/wiki/Ciclo_del_nitr%C3%B3geno.
[31] Craine, J.M., Brookshire, E.N.J., Cramer, M.D., Hasselquist, N.J., Koba, K., Marin-Spiotta, E. y Wang, L., 2015. Ecological interpretations of nitrogen isotope ratios of terrestrial plants and soils. Plant Soil, 396, 1–26. https://link.springer.com/article/10.1007/s11104-015-2542-1.
[32] Garvin, J., Buick, R., Anbar, A.D., Arnold, G.L. y Kaufman, A.J., 2009. Isotopic evidence for an aerobic nitrogen cycle in the latest Archean. Science, 323 (5917), 1045-1048. http://science.sciencemag.org/content/323/5917/1045.
[33] Reinhard, C.T. y otros, 2017. Evolution of the global phosphorus cycle. Nature, 541, 386-389. www.nature.com/articles/nature20772.
[34] Lessner, D.J., 2009. Methanogenesis biochemistry. Encyclopedia of Life Sciences. https://onlinelibrary.wiley.com/doi/full/10.1002/9780470015902.a0000573.pub2.
[35] Cameron, V., Vance, D., Archer, C. y House, C.H., 2009. A biomarker based on the stable isotopes of nickel. Proceedings of the National Academy of Sciences, 106 (27), 10944-10948. www.pnas.org/content/pnas/106/27/10944.full.pdf.
[36] Ver: https://en.wikipedia.org/wiki/Biomarker.
[37] Ver: https://en.wikipedia.org/wiki/Sterane; www-eaps.mit.edu/geobiology/biomarkers/steroids.html; https://es.wikipedia.org/wiki/Ciclopentanoperhidrofenantreno.
[38] Cavalier-Smith, T., Brasier, M. y Embley, T.M., 2006. Introduction: how and when did microbes change the world? Philosophical Transactions of the Royal Society B, 361, 845-850. http://rstb.royalsocietypublishing.org/content/361/1470/845.
[39] Summons, R.E., Bradley, A.S., Jahnke, L.L. y Waldbauer, J.R., 2006. Steroids, triterpenoids and molecular oxygen. Philosophical Transactions of the Royal Society B, 361, 951-968. http://rstb.royalsocietypublishing.org/content/361/1470/951. Fischer, W.W., 2008. Life before the rise of oxygen. Nature, 455, 1051-1052. www.nature.com/articles/4551051a.
[40] Buick, R., Rasmussen, B. y Krapez, B., 1998. Archean oil: evidence for extensive hydrocarbon generation and migration 2.5-3.5 Ga. AAPG Bulletin, 82 (1), 50-69. www.aapg.org.
[41] Woese, C.R., Kandler, O. y Wheelis, M.L., 1990. Towards a natural system of organisms: proposal for the domains Archaea, Bacteria, and Eucarya. Proceedings of the National Academy of Sciences, 87 (12), 4576-4579. www.pnas.org/content/87/12/4576.long.
[42] Ver: https://es.wikipedia.org/wiki/Dominio_(biolog%C3%ADa).
[43] Ver: https://es.wikipedia.org/wiki/Archaea.
[44] Kelly, S., Wickstead, B. y Gull, K., 2010. Archaeal phylogenomics provides evidence in support of a methanogenic origin of the Archaea and a thaumarchaeal origin for the eukaryotes. Proceedings of the Royal Society B, 278, 1009-1018. http://rspb.royalsocietypublishing.org/content/royprsb/278/1708/1009.full.pdf. Sarmiento, F., Mrázek, J. y Whitman, W.B., 2013. Genome-scale analysis of gene function in the hydrogenotrophic methanogenic archaeon Methanococcus maripaludis. Proceedings of the National Academy of Sciences, 110 (12), 4726-4731. www.pnas.org/content/110/12/4726.
[45] Chapelle, F.H., O’Neill, K., Bradley, P.M., Methé, B.A., Ciufo, S.A., Knobel, L.L. y Lovley, D.R., 2002. A hydrogen-based subsurface microbial community dominated by methanogens. Nature, 415, 312-315. www.nature.com/articles/415312a.
[46] Ver: https://es.wikipedia.org/wiki/Cyanobacteria.
[47] Gribaldo, S. y Brochier-Armanet, C., 2006. The origin and evolution of Archaea: a state of the art. Philosophical Transactions of the Royal Society B, 361, 1007-1022. http://rstb.royalsocietypublishing.org/content/361/1470/1007.
[48] Zhang, C.L., Pearson, A., Li, Y.-L., Mills, G. y Wiegel, J., 2006. Thermophilic temperature optimum for crenarchaeol synthesis and its implication for Archaeal evolution. Applied and Environmental Microbiology, 72 (6), 4419-4422. http://aem.asm.org/content/72/6/4419.full.pdf. Huang, Z., Hedlund, B.P., Wiegel, J., Zhou, J. y Zhang, C.L., 2007. Molecular phylogeny of uncultivated Crenarchaeota in Great Basin hot springs of moderately elevated temperature. Geomicrobiology Journal, 24 (6), 535-542. www.tandfonline.com/doi/abs/10.1080/01490450701572523.
[49] Ueno, Y., Yamada, K., Yoshida, N., Maruyama, S. e Isozaki, Y., 2006. Evidence from fluid inclusions for microbial methanogenesis in the early Archaean era. Nature, 440, 516-519. www.nature.com/articles/nature04584.
[50] Nelson-Sathi, S. y otros, 2015. Origins of major archaeal clades correspond to gene acquisitions from bacteria. Nature, 517, 77-80. www.nature.com/articles/nature13805.
[51] Williams, T.A., Szöllősi, G.J., Spang, A., Foster, P.G., Heaps, S.E., Boussau, B., Ettema, T.J.G. y Embley, T.M., 2017. Integrative modeling of gene and genome evolution roots the archaeal tree of life. Proceedings of the National Academy of Sciences, 114 (23), E4602-E4611. www.pnas.org/content/pnas/114/23/E4602.full.pdf.
[52] Rice, G. y otros, 2004. The structure of a thermophilic archaeal virus shows a double-stranded DNA viral capsid type that spans all domains of life. Proceedings of the National Academy of Sciences, 101 (20), 7716-7720. www.pnas.org/content/pnas/101/20/7716.full.pdf. Benson, S.D., Bamford, J.K.H., Bamford, D.H. y Burnett, R.M., 2004. Does common architecture hypothesis reveal a viral lineage spanning all three domains of life? Molecular Cell, 16, 673-685. www.cell.com/molecular-cell/pdf/S1097-2765(04)00709-9.pdf. Laidler, J.R. y Stedman, K.M., 2010. Virus silicification under simulated hot spring conditions. Astrobiology, 10 (6), 569-576. www.liebertpub.com/doi/abs/10.1089/ast.2010.0463.
[53] Wang, M., Yafremava, L.S., Caetano-Anollés, D., Mittenthal, J.E. y Caetano-Anollés, G., 2007. Reductive evolution of architectural repertoires in proteomes and the birth of the tripartite world. Genome Research, 17, 1572-1585. https://genome.cshlp.org/content/17/11/1572.full.pdf. Zeldovich, K.B., Chen, P., Shakhnovich, B.E. y Shakhnovich, E.I., 2007. A first-principles model of early evolution: emergence of gene families, species, and preferred protein folds. PLoS Computational Biology, 3 (7), e139. http://journals.plos.org/ploscompbiol/article?id=10.1371/journal.pcbi.0030139.
[54] Fournier, G.P. y Alm, E.J., 2015. Ancestral reconstruction of a pre-LUCA Aminoacyl-tRNA synthetase ancestor supports the late addition of Trp to the genetic code. Journal of Molecular Evolution, 80 (3-4), 171-185. https://link.springer.com/article/10.1007/s00239-015-9672-1.
[55] Busch, F., Rajendran, C., Heyn, K., Schlee, S., Merkl, R. y Sterner, R., 2016. Ancestral tryptophan synthase reveals functional sophistication of primordial enzyme complexes, Cell Chemical Biology, 23, 1-7. www.cell.com/cell-chemical-biology/pdf/S2451-9456(16)30162-3.pdf.
[56] Singer, E., 2015. How structure arose in the primordial soup. Quanta Magazine. www.quantamagazine.org/20150416howstructureevolvedintheprimordialsoup.
[57] Lewis Jr., C.A., Crayle, J., Zhou, S., Swanstrom, R. y Wolfenden, R., 2016. Cytosine deamination and the precipitous decline of spontaneous mutation during Earth’s history. Proceedings of the National Academy of Sciences, 113 (29), 8194-8199. www.pnas.org/content/113/29/8194.
[58] Rousseau, B.J.G., Shafei, S., Migliore, A., Stanley, R.J. y Beratan, D.N., 2018. Determinants of photolyase’s DNA repair mechanism in mesophiles and extremophiles. Journal American Chemical Society, 140 (8), 2853-2861. https://pubs.acs.org/doi/10.1021/jacs.7b11926.
[59] Mohr, G., Ghanem, E. y Lambowitz, A.M., 2010. Mechanisms used for genomic proliferation by thermophilic group II introns. PLoS Biology, 8 (6), e1000391. http://journals.plos.org/plosbiology/article?id=10.1371/journal.pbio.1000391. Ver también: https://es.wikipedia.org/wiki/Intr%C3%B3n.
[60] Ver: https://en.wikipedia.org/wiki/Methanogenesis.
[61] Czaja, A.D., Beukes, N.J. y Osterhout, J.T., 2016. Sulfur-oxidizing bacteria prior to the Great Oxidation Event from the 2.52 Ga Gamohaan Formation of South Africa. Geology, 44 (12), 983-986. http://geology.gsapubs.org/content/44/12/983.full.pdf.
[62] Braakman, R. y Smith, E., 2012. The emergence and early evolution of biological carbon-fixation. PLoS Computational Biology, 8 (4), e1002455. http://journals.plos.org/ploscompbiol/article?id=10.1371/journal.pcbi.1002455.
[63] Ver: https://es.wikipedia.org/wiki/%C3%9Altimo_antepasado_com%C3%BAn_universal#cite_note-3.
[64] Martin, W., 1999. Mosaic bacterial chromosomes: a challenge en route to a tree of genomes. BioEssays, 21 (2), 99-104. https://onlinelibrary.wiley.com/doi/abs/10.1002/%28SICI%291521-1878%28199902%2921%3A2%3C99%3A%3AAID-BIES3%3E3.0.CO%3B2-B. Doolittle, W.F., 2000. Uprooting the tree of life. Scientific American, 282 (2), 90-95. https://www.nature.com/scientificamerican/journal/v282/n2/pdf/scientificamerican0200-90.pdf.
[65] Poole, A., 2009. Mi nombre es LUCA – El último ancestro universal común. http://actionbioscience.org/esp/nuevas-fronteras/poolepaper.html.
[66] Weiss, M.C., Sousa, F.L., Mrnjavac, N., Neukirchen, S., Roettger, M., Nelson-Sathi, S. y Martin, W.F., 2016. The physiology and habitat of the last universal common ancestor. Nature Microbiology, 1, 16116. www.nature.com/articles/nmicrobiol2016116.
[67] Boussau, B., Blanquart, S., Necsulea, A., Lartillot, N. y Gouy, M., 2008. Parallel adaptations to high temperatures in the Archaean eon. Nature, 456, 942-945. www.nature.com/articles/nature07393.
[68] Wang y otros, 2007. Ver nota 53.
[69] Caforio, A. y otros, 2018. Converting Escherichia coli into an archaebacterium with a hybrid heterochiral membrane. Proceedings of the National Academy of Sciences, 115 (14), 3704-3709. www.pnas.org/content/115/14/3704.
[70] Doolittle, 2000. Ver nota 64.
[71] Doolittle, 2000. Ver nota 64. Glansdorff, N., Xu, Y. y Labedan, B., 2008. The Last Universal Common Ancestor: emergence, constitution and genetic legacy of an elusive forerunner. Biology Direct, 3, 29. https://biologydirect.biomedcentral.com/track/pdf/10.1186/1745-6150-3-29.
[72] Marin, J., Battistuzzi, F.U., Brown, A.C. y Hedges, S.B., 2016. The timetree of prokaryotes: new insights into their evolution and speciation. Molecular Biology and Evolution, 34 (2), 437-446. https://academic.oup.com/mbe/article/34/2/437/2740734.

Hola Pieter, soy un geólogo venezolano que trabaje por muchos años en el área de mineralogía y petrología en el ministerio de Energía y Minas y te conocí en una excursión que organizo la Soc Vzlana de Geólogos al estado Falcón. Recientemente conseguí tu blog y me pareció muy interesante, por lo cual te felicito por tu dedicación desinteresada por llevar a tanta gente tu visión del origen de la vida desde la geología. Pieter, me gustaría leer algo sobre la extinción de las especies a través del tiempo geológico, si conoces de alguna bibliografía sobre este tema y especialmente si esta en español, te agradecería que me enviaras las referencias
Saludos
Colega y Amigo
Oswaldo Cisneros G
Me gustaMe gusta
Hola Oswaldo, gracias por tus palabras, y ¡qué memoria tienes! Tengo previsto escribir acerca de las grandes extinciones después de la actual serie de entradas sobre el desarrollo de la vida. Pero no será pronto… Como punto de partida de un estudio sobre la extinción puedes utilizar la entrada «extinción» de Wikipedia, y seguir los vínculos que allí se encuentran. ¡Éxito! Un gran saludo, Pieter
Me gustaMe gusta