
El día que la vida apareció en la tierra debe haber lucido como un día igual a todos los demás. Un sol joven iluminaba un planeta inhóspito, dominado por colores rojizos y grisáceos en el mar, la tierra y el cielo, azotado por impactos de meteoritos y volcanismo (ver la primera entrega de esta serie, Los albores de la vida). Pero en un lugar escondido, en algún cuerpo de agua que a primera vista era como muchos otros lugares del planeta, ocurrieron procesos químicos que dieron origen a conjuntos de moléculas capaces de mantenerse y multiplicarse: la primera vida.
Para entender la vida es preciso estudiar sus orígenes, que al fin y al cabo son también los del género humano. Además, si queremos saber si puede haber surgido alguna forma de vida en otros cuerpos celestes, tenemos que empezar por la única que conocemos: la terrestre.
En la entrega anterior vimos cuándo aparecieron los primeros organismos terrestres. Hace unos 4 mil millones de años, ciertas moléculas complejas comenzaron a hacer copias de sí mismas, un evento que marcó el comienzo de la vida en la tierra. Unos cientos de millones de años más tarde, los organismos primitivos comenzaron a diferenciarse en las distintas ramas que componen el árbol de la vida. Entre esos dos eventos seminales surgieron algunas de las mayores innovaciones biológicas: la célula, el código genético y un sistema de energía para alimentarlo todo. Los tres son esenciales para la vida tal como la conocemos; sin embargo, los científicos aun están investigando cómo surgieron estas innovaciones[1]. Veamos qué sabemos hoy día de cómo puede haber sido el origen de la vida.
En la entrega del 19 mayo 2018 (¿Qué es la vida?) vimos cuál es la definición de la vida que ahora tiene mayor aceptación. Según esta definición, la vida es:
Un sistema químico autosustentable capaz de experimentar evolución darwiniana.
Para que pueda cumplir con esta definición, un organismo primordial necesita tener las siguientes características:
- Tener un envoltorio, para que pueda ser considerado un sistema separado de su entorno. Este contenedor es, desde los primeros organismos que conocemos, la membrana externa de la célula.
- Tener un mecanismo para generar energía, para que sea autosustentable. Esto se hace mediante la ingesta de alimentos de afuera del organismo, que son convertidos en energía y desechos; esto es lo que se llama el metabolismo.
- Contar con un conjunto de instrucciones de cómo ensamblarse, para que sea autosustentable y para que pueda actuar la evolución darwiniana. Este debe ser transmisible a los descendientes. El material genético cumple esta función, por lo menos desde los tiempos del LUCA, el último ancestro común de todos los organismos actuales (ver la entrega anterior).
- Ser capaz de dividirse, para que pueda actuar la evolución darwiniana. Esto permite la multiplicación de los organismos y la permanencia de la especie.
Todas estas características son indispensables para que se pueda hablar de vida. Pero, ¿cómo aparecieron?, y, ¿cuál apareció primero?
Para reconstruir cómo pueden haber sido los primeros organismos (e intentar construir un organismo en el laboratorio), existen dos enfoques: uno, “top-down”, o sea, desde arriba (lo complejo) hacia abajo (las bases), y otro, “bottom-up”, en sentido opuesto. El primer enfoque deconstruye (deshace) los más simples organismos unicelulares actuales para ver cuáles son los elementos indispensables para que los organismos puedan funcionar; estos suelen dar por elemento clave el ARN (como veremos a continuación), no el ADN. Pero no es probable que el ARN, una molécula compleja, haya existido desde los albores de la vida, y por tanto pudiera tener más mérito el enfoque bottom-up, que empieza por los compuestos y procesos que pueden haber jugado un papel en la formación de los primeros organismos[2]. Sigamos, por tanto, este segundo enfoque.
El agua
Se considera que los primeros organismos terrestres se originaron en un medio acuático. El agua es un diluyente ideal para muchas reacciones químicas. Existen otros diluyentes, tales como el metano en la gélida luna Titán de Saturno[3], y veremos más adelante que en nuestro planeta pudo haber existido otros diluyentes también, que pueden haber jugado un papel en la primera vida. Pero el agua es el medio más abundante, y por tanto el más obvio para el desarrollo de la vida. Tal como veremos en la próxima entrega, no se sabe si los primeros organismos se formaron en el mar o en lagos en un ambiente terrestre – pero en algún medio acuático debe haber sido.
El contenedor
En esta entrega estaremos hablando mucho de las primeras moléculas de la vida. Pero es importante recordarnos que éstas no se desarrollaron flotando en el medio del mar o algún otro cuerpo acuático: deben haber estado ubicadas dentro de algún tipo de contenedor. El contenedor es necesario para concentrar los compuestos que van a dar origen a las moléculas que componen el organismo. Además, sin contenedor no se puede conservar la integridad del organismo, así que se supone que ya en una fase muy temprana del desarrollo de la vida debe haber aparecido algún tipo de membrana que funcionara como envoltorio.
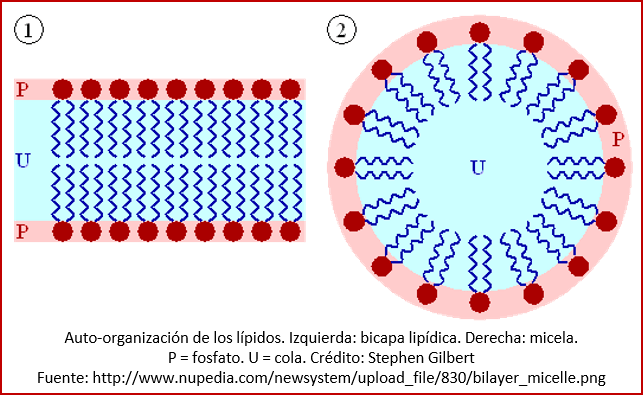
Todos los organismos conocidos tienen células cuyas membranas son formadas por fosfolípidos, un tipo de grasas. Los organismos actuales tienen membranas relativamente complejas, con una diferenciación entre las membranas de las arqueas y las de los demás organismos (ver la entrada anterior de este blog); pero se supone que los primeros organismos tuvieron una membrana fosfolipídica más simple[4]. Los fosfolípidos consisten en un grupo fosfato con una cola compuesta de un alcohol y dos ácidos grasos. Debido a su carácter hidrofóbico (o sea, experimentan una repulsión al agua), los fosfolípidos se unen fácilmente, formando membranas bicapa o micelas (mini-vesículas; ver imagen)[5]. Los fosfolípidos se forman mediante procesos abióticos (es decir, que no dependen de ningún proceso biológico) y, por tanto, son candidatos obvios para haber formado las membranas de las primeras células[6]. (Pero también se ha propuesto que ciertos polipéptidos formados por aminoácidos pueden haber servido como membranas primordiales[7].)
Una membrana celular debe ser lo suficientemente permeable para que pueda ocurrir un intercambio de compuestos específicos entre la célula y su entorno. Células modernas tienen mecanismos complejos para esto. Sin embargo, se ha demostrado que también las membranas fosfolipídicas más sencillas son capaces de dejar pasar ciertos compuestos relevantes para la célula[8].
Las vesículas de fosfolípidos crecen sencillamente, mediante la incorporación de moléculas de lípidos y de micelas enteras[9]. Además, tienen otra gran ventaja: después de haber alcanzado cierto tamaño pueden dividirse fácilmente, lo que es un proceso fisicoquímico que no requiere de la acción de biomoléculas (aunque es posible que sea facilitado por la replicación del material genético[10]), sino que depende de deformaciones causadas por las tensiones entre la superficie y el interior la célula[11]. De esta manera, las primeras células pueden haberse dividido, dando origen a la replicación del organismo[12]. En las primeras células, las membranas eran presumiblemente menos rígidas que las de las bacterias actuales, lo que permitía una división de las células mediante un proceso de expulsión de vesículas “hijas” de la célula madre[13].
Dentro de una célula pueden formarse pequeñas vesículas con su propia membrana. Estas suelen atrapar una cantidad grande de moléculas presentes dentro de la célula, y en algunos casos estas moléculas tienen una cantidad y concentración suficiente para la formación de compuestos complejos tales como proteínas[14].
Hasta ahora hemos hablado de membranas formadas por compuestos orgánicos. Sin embargo, también se ha observado, en ambientes específicos tales como aguas muy alcalinas, ricas en sílice, la formación de membranas inorgánicas compuestas por hidratos de metales y sílice. Estas membranas, aparte de servir como envoltorio para las primeras células, pueden haber actuado como catalizadores para la formación de compuestos orgánicos prebióticos, o sea, precursores de las moléculas biológicas[15]. Otros polímeros (moléculas largas formadas por una encadenación de compuestos más sencillos), tales como el polietilenglicol y dextrano, pueden haber formado compartimentos dentro de un medio acuático, en las que biomoléculas como el ARN pueden haber alcanzado concentraciones suficientemente altas para empezar a participar en reacciones químicas[16].
Como alternativas a una membrana como envoltorio de los primeros organismos, se ha propuesto varios tipos de cavidades en minerales o rocas en el fondo marino o lacustre. Ciertos sulfuros de hierro, por ejemplo, tienen una estructura en la forma de un panal de abejas; en los huecos dentro de esta estructura pueden haberse desarrollado los primeros organismos, utilizando el mineral como envoltorio, en lugar de una membrana[17]. Tales sulfuros de hierro pueden formarse en los alrededores de fuentes de agua caliente en el fondo marino, que son lugares en los que, según una teoría bastante difundida, puede haberse desarrollado la primera vida terrestre (ver la siguiente entrega de este blog). En estos ambientes pueden haberse precipitado también sulfuros de zinc, que además han sido propuestos como cuna de los primeros organismos, ya que, aparte de servir como envoltorio, pueden haber servido como catalizadores y ayudado en la reducción de dióxido de carbono, para así formar los componentes de las primeras moléculas biológicas[18]. Abajo veremos más ejemplos de cómo minerales pueden haber ayudado en el desarrollo de los primeros organismos..
Expresión genética
Antes de entrar en el tema del origen de la vida, es preciso hacer un breve repaso de cómo funciona, en las células de los organismos actuales, la traducción de la información genética en instrucciones acerca del funcionamiento, crecimiento y reproducción de las células. Este proceso se denomina la expresión genética; es el “dogma central” de la biología molecular. Los principales compuestos involucrados son el ADN y el ARN.
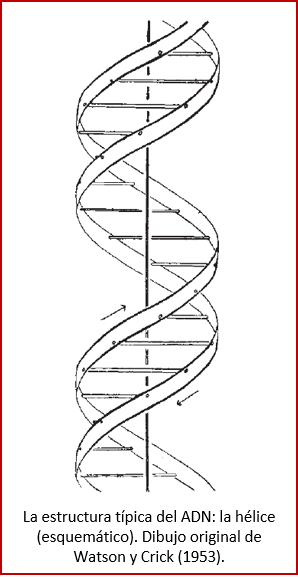
El ADN, o sea, ácido desoxirribonucleico, contiene las instrucciones genéticas usadas en el desarrollo y funcionamiento de todos los organismos vivos. También es responsable de la transmisión hereditaria. La función principal de la molécula de ADN es el almacenamiento a largo plazo de información para construir otros componentes de las células, como las proteínas (los compuestos que llevan a cabo las labores de construir, mantener y reproducir las células) y las moléculas de ARN. Los segmentos de ADN que llevan esta información genética son llamados genes. El ADN es una molécula bicatenaria, es decir, está formada por dos cadenas, una frente la otra, y desde ya hace más de sesenta años sabemos que su estructura es la de una hélice[19] (ver figura). Desde el punto de vista químico, el ADN es un polímero (encadenación) de nucleótidos. Cada nucleótido, a su vez, está formado por un azúcar (la ribosa), una nucleobase (que puede ser adenina (A), timina (T), citosina (C) o guanina (G)), y un fosfato[20].
Por otro lado, el ARN, o sea ácido ribonucleico, es parecido al ADN, pero comúnmente es formado por una sola cadena de ribonucleótidos. Es la molécula que dirige las etapas intermedias de la síntesis proteica; el ADN no puede actuar solo, y se vale del ARN para transferir esta información vital durante la producción de proteínas. El ARN es más versátil que el ADN: varios tipos de ARN regulan la expresión génica, mientras que otros (llamados ribozimas) funcionan como enzimas; actúan como catalizadores, facilitando reacciones químicas en la célula[21].
Los pasos por seguir para realizar la expresión genética, o sea, traducir la información genética en proteínas, son los siguientes (ver figura):
- Transcripción. La transcripción del ADN es el primer paso de la expresión genética. Durante este proceso, ciertas secuencias de ADN son copiadas a ARN mediante una enzima llamada ARN polimerasa la cual sintetiza un ARN mensajero (ARNm) que mantiene la información de la secuencia del ADN[22].
- Traducción. La traducción ocurre por lo general en el citoplasma de la célula (no en el núcleo, si la célula lo contiene), donde se encuentran los ribosomas. Los ribosomas consisten en ARNr (ARN ribosomal) y proteínas. El ribosoma y las moléculas de ARNt (ARN transferente) traducen el código del ARNm para producir proteínas. En la traducción, el ARNm se decodifica para generar una cadena específica de aminoácidos y así formar una proteína[23]. El ribosoma es una biomolécula muy antigua, cuyos orígenes, tal como veremos abajo, se remontan al amanecer de la vida[24]. Ya en una fase muy temprana se estableció en el ribosoma un mecanismo para codificar en el material genético cada uno de los veinte aminoácidos que son piezas clave de las que se componen la mayoría de las biomoléculas[25].

Es un proceso complejo, que no puede haber aparecido de la nada cuando se desarrollaron los primeros organismos en la tierra primordial. Por tanto, la gran pregunta es, ¿cómo puede haberse desarrollado, a partir de compuestos abióticos (o sea, moléculas formadas por procesos no biológicos), el mecanismo que permite registrar la información para el funcionamiento de la célula y leerla para que se lleven a cabo los procesos respectivos? De esto estaremos hablando en la presente, y la próxima, entrega.
Las piezas prebióticas
Otra gran pregunta, relacionada a la anterior, es, ¿cómo pueden haberse formado, en los primeros organismos, las biomoléculas complejas actuales a partir de los compuestos simples del mundo inanimado? Las moléculas que actualmente se encuentran dentro de las células de los organismos suelen ser complejas, tanto en términos de composición química como de la manera en la cual se han plegado. Los procesos de formación de estas moléculas son complicados, y – así como la gran mayoría de las reacciones químicas que ocurren dentro de las células – requieren de la ayuda de catalizadores (o sea, enzimas, que son un tipo de proteínas).
Respuestas exactas a estas preguntas aun no las tenemos, pero parece que hubo, entre las moléculas orgánicas (moléculas con carbono e hidrógeno), moléculas prebióticas que sirvieron como base para la elaboración de las moléculas biológicas que se encuentran en los organismos. Estas moléculas prebióticas se forman mediante procesos abióticos. Por ejemplo, vimos en la entrega de este blog del 26 agosto 2017 (El preludio de la vida (6): Semillas universales) que hasta en el espacio tales moléculas, incluyendo algunos aminoácidos, se están formando. Esta facilidad de formar aminoácidos es probablemente una de las razones por que estos ácidos son las principales piezas de las que se componen muchas biomoléculas, específicamente las proteínas.
En la tierra primordial, las moléculas prebióticas se pudieron formar también (ver la entrega del 26 agosto 2017, arriba mencionada), aportando los compuestos necesarios para elaborar las piezas de las que se componen las biomoléculas. En el caso del ADN y el ARN, se ha identificado moléculas prebióticas que pueden haber servido para armar tanto las nucleobases como la ribosa y el fosfato. Los dos principales grupos de nucleobases son las purinas y las pirimidinas; las letras A y G del ADN son purinas, y las letras C y T son pirimidinas[26]. Se ha encontrado un posible precursor prebiótico de ambos grupos: un azúcar, que puede haberse modificado tanto en una pirimidina como en una purina[27]. La ribosa puede haberse formado de distintas maneras (en general, los azúcares se forman fácilmente), aunque sólo es estable bajo ciertas condiciones, por ejemplo en la presencia de borato[28]. El fosfato, que no es una molécula orgánica, puede haberse formado a partir de la disolución, en ambientes calientes (volcánicos o hidrotermales), de minerales contentivos de fósforo, o puede haber provenido de minerales fosfáticos en meteoritos[29]. También se determinó que es posible formar moléculas precursoras del ARN a partir de una solución acuosa con altas concentraciones de cianuro de hidrógeno y sulfuro de hidrógeno, bajo la incidencia de rayos ultravioletas – ingredientes que probablemente estuvieron presentes en la tierra primordial[30]. Así que no es tan difícil formar las piezas de las que se componen los nucleótidos. Sin embargo, tal como veremos abajo, juntarlas para formar los nucleótidos y después las cadenas del ARN y ADN, es otro cantar.
De particular relevancia para la vida es la formación de los péptidos (los “hermanos menores” de las proteínas; ambos consisten en cadenas de aminoácidos), que realizan, conjuntamente a las proteínas, gran parte de las labores dentro de las células[31]. Se ha logrado formar péptidos en el laboratorio a partir de aminoácidos en condiciones parecidas a las que pueden haber existido en la tierra primordial[32]. Uno de estos péptidos recreados en el laboratorio es capaz de facilitar el ciclo metabólico de las células[33].
Los aminoácidos son la base de muchos compuestos en la célula. Hoy día existen veinte, pero se sabe que inicialmente hubo menos, y que los aminoácidos más sencillos se pueden formar de manera abiótica (ver la entrega del 26 agosto 2017). Recientemente se determinó, a partir de vestigios en la estructura de moléculas del ARNt (que transportan aminoácidos para el ensamblaje de proteínas), que inicialmente los organismos utilizaban sólo dos aminoácidos – lo que implica que los mecanismos celulares para la producción de proteínas pueden haber sido más sencillos de lo que son hoy día[34].
También se ha encontrado una molécula que, en agua a temperatura ambiente, es capaz de promover la producción de no sólo péptidos, sino también lípidos (que forman las membranas externas de las células) e inclusive nucleótidos (de los que se componen el ADN y el ARN). Se trata del diaminofosfato, que, al aportar fósforo en la forma de fosfato, es clave en la elaboración de las clases de compuestos mencionadas y por tanto pudiera resultar fundamental en el entendimiento de cómo pueden haberse formado los primeros organismos[35].
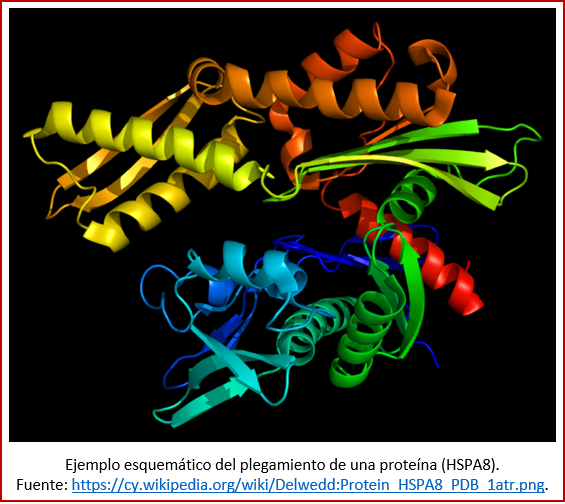
Un factor que hace difícil la elaboración de las enzimas es su estructura globular: las enzimas, así como las proteínas en general, están plegadas de manera complicada, y única para cada compuesto (ver imagen). Estas estructuras son difíciles de sintetizar a partir de procesos prebióticos. Sin embargo, en ciertas condiciones específicas, tales como un ambiente acuático muy salino, se ha logrado que se plegaran unas proteínas compuestas por aminoácidos sencillos[36]. Alternativamente, puede haber habido una fase intermedia de enzimas híper-ramificadas en lugar de globulares, que sí pueden formarse mediante procesos prebióticos y que funcionan de manera semejante a las enzimas globulares[37].
Cabe destacar que, de todos los compuestos orgánicos posibles, la vida terrestre sólo está utilizando una pequeña parte. ¿Es casualidad, o hubo una selección natural de compuestos orgánicos cuando se formó la primera vida y quedaron descartados aquellos compuestos que no servían? Este es otro asunto que queda por aclarar[38]. Lo que sí podemos suponer, es que hubo muchos compuestos más de los que encontramos ahora en la naturaleza, y que algunos de estos pueden haber sido importantes para el desarrollo de la primera vida[39].
¿Substrato catalizador?
Para la formación de moléculas complejas tales como las biológicas, se ha propuesto que pueden haber servido como catalizadores ciertos minerales. Los catalizadores (compuestos que ayudan las reacciones químicas) son necesarios para la elaboración de la gran mayoría de las moléculas biológicas. Si las enzimas, que actualmente sirven como catalizadores en las células de los organismos, todavía no existían en los albores de la vida, debe haber habido otros catalizadores, y ciertos minerales pueden haber cumplido esta función.
En la búsqueda de minerales que pueden haber ayudado en la formación de los primeros organismos es importante darnos cuenta de que no todos los minerales que ocurren hoy en día, existieron en la tierra primordial. Al no haber oxígeno en la atmósfera, tampoco había muchos minerales contentivos de este elemento. Afortunadamente eran comunes las arcillas y los sulfuros, que según muchos modelos eran importantes para que se pudieran desarrollar las primeras formas de vida[40].
Ciertas arcillas pueden servir como substrato para la formación del ARN (ver imagen) [41], y un mineral como la molibdenita puede haber servido como substrato para la formación de biomoléculas tales como la adenina, que es una de las bases del ADN[42]. Se determinó experimentalmente que el grafito (una forma de carbono) y la molibdenita, que forman cristales con superficies planas, pueden adsorber (retener en su superficie) moléculas orgánicas, tales como la adenina y la guanina, que forman estructuras regulares bidimensionales en las superficies de los cristales[43].

El diamante (una forma cristalina de carbono), si se expone al hidrógeno, desarrolla una película de agua superficial en la que pudieran formarse moléculas orgánicas, utilizando al carbono del diamante como materia prima; además, una vez formadas, las películas de agua mejoran el funcionamiento de los organismos que en ellas se encuentran[44].
Aparte de las arcillas, se ha determinado que las micas pueden haber funcionado como cuna para los primeros organismos. Estos minerales consisten en un apilamiento de láminas delgadas, entre las que pueden haberse formado las moléculas complejas de la vida. Pero, además, las micas contienen cierta cantidad de agua, que es necesaria para que las reacciones químicas se puedan dar, y proporcionan un ambiente protegido, aislando las moléculas del mundo externo y sirviendo como envoltorio de la proto-célula, en lugar de una membrana[45].
Otro material del cual se ha propuesto que puede haber servido como substrato para la primera vida, es roca porosa. Tales rocas proporcionan un ambiente protegido en el que puede mantenerse, y haberse formado, la vida[46]. En particular se puede mencionar la piedra pómez: una roca volcánica altamente porosa, tanto así que es capaz de flotar en el agua. Esta roca tiene muchos minerales y, al estar expuesta a la intemperie flotando en el agua, era un substrato ideal para la vida primordial ya que pudo hacer posible una gran cantidad de reacciones químicas y así ayudar en la generación de las primeras moléculas biológicas[47]. Otra roca de interés es la serpentinita (una roca volcánica alterada por procesos hidrotermales), que, al reaccionar con agua marina, puede liberar hidrógeno y otros elementos relevantes para la formación y el funcionamiento de los primeros organismos[48].
Otro tipo de rocas que pueden haber protegido la primera vida y facilitado su aparición, son aquellas rocas fracturadas y recristalizadas como resultado del impacto de un meteorito. Actualmente estas rocas son raras, pero hace alrededor de 4000 millones de años, cuando empezó a formarse la vida terrestre, eran comunes a causa del bombardeo intenso de meteoritos que en aquellos tiempos sufrió nuestro planeta. Estas rocas impactadas contienen muchos microporos y, por los daños en los cristales debido al impacto, protegen a los organismos en los poros de los rayos ultravioletas que los pueden dañar y hasta matar[49]. Esta capacidad protectora ha sido comprobada en experimentos[50]. Además, las temperaturas elevadas en la zona del impacto (que pueden perdurar por decenas de miles de años[51]), combinadas con la circulación de aguas calientes, hidrotermales, pueden haber fomentado el desarrollo de la vida. En efecto, se ha encontrado formas tubulares microscópicas en rocas vitrificadas por el impacto de un meteorito, que han sido interpretado como trazas de organismos que vivieron en la superficie de estas rocas después del impacto[52]. Además, tales rocas porosas con poros alargados pueden haber facilitado, en la presencia de peróxido de hidrógeno, la formación de las primeras moléculas orgánicas alargadas[53].
Tal vez no sea una coincidencia que los primeros organismos terrestres hayan aparecido en un tiempo que había muchos impactos de meteoritos. Además de los beneficios de las rocas impactadas como cuna de la vida, tenemos que tomar en cuenta los compuestos orgánicos traídos por los meteoritos del espacio, los cuales pueden haber servido como base para las moléculas biológicas necesarias para la primera vida[54].
En lugar de las rocas afectadas por los impactos de los meteoritos, puede haber sido la intensa actividad volcánica, causada por estos impactos, la que facilitó la aparición de la vida. Según una hipótesis que enfatiza el papel de las rocas y los minerales en el desarrollo de los primeros organismos, se acumularon minerales provenientes de cenizas volcánicas y flujos de lava en charcas a los pies de los volcanes. En estas charcas efímeras, los minerales (que incluían feldespatos y zeolitas) estaban expuestos a secado e inundación intermitente, lo que fomentaba la adsorción y los procesos catalíticos. Algunas superficies ricas en sílice favorecían la adsorción de moléculas orgánicas, incluidos los aminoácidos, que se producían mediante rayos en las nubes volcánicas. Catálisis luego promovió la polimerización para generar moléculas más complejas. La disolución de feldespatos creó un entramado de poros, que pudieran haber actuado como paredes celulares temporales, mientras que el fósforo liberado de los feldespatos disueltos estaba disponible para la formación de las moléculas que proveen la energía a los organismos[55].
Una vez aparecidas las membranas lipídicas de las primeras células, no necesariamente terminó la importancia de las arcillas. Se ha sugerido que partículas de arcillas, que a menudo quedan incorporadas en vesículas de lípidos, pueden haber actuado de vehículo para las biomoléculas, adsorbiendo moléculas tales como el ARN[56].
La estrecha relación que, según estas teorías, existió entre los primeros organismos por un lado y rocas y minerales por otro, pudiera explicar por qué son altas las concentraciones de ciertos elementos, aun hoy día, en las células de los organismos: las concentraciones relativamente altas de potasio y zinc pudieran reflejar la mineralogía de las rocas en las que, posiblemente, se formaron los primeros organismos[57].
Oparin y la sopa primordial
Una de las primeras teorías acerca de cómo la vida puede haber aparecido en la tierra, es la del bioquímico ruso Aleksandr Oparin (1894-1980), quien en 1924 afirmó que la vida en la tierra se desarrolló a través de cambios químicos graduales de moléculas orgánicas, en una “sopa primordial” que probablemente existió en la tierra hace unos cuatro mil millones de años. En su opinión, la compleja combinación de moléculas sin vida, dentro de pequeños glóbulos aceitosos, pudiera alcanzar las propiedades de la vida: autorreplicación, selección y evolución[58]. Estas ideas recibieron apoyo de los ahora famosos experimentos de Stanley Miller de los años ’50 [59], que indicaron que ciertas moléculas orgánicas, relevantes como base para los compuestos biológicos, pudieron haberse formado bajo condiciones parecidas a las que prevalecían cuando la tierra estaba recién formada.
Sin embargo, la teoría de Oparin fue descartada cuando se empezó a conocer los portadores de la información genética: el ADN. Se consideró que, puesto que esta biomolécula es capaz de autorreplicarse, quedó resuelto el enigma del origen de la vida sin tener que recurrir a los glóbulos de Oparin. Pero abajo veremos que, ante la dificultad de crear a partir de compuestos prebióticos los biopolímeros informacionales (o sea, el ADN y el ARN), algunas ideas de Oparin han vuelto a estar siendo investigadas.
El mundo ADN
El ADN, al ser el portador de toda la información genética, es fundamental para la vida como la conocemos. Que haya sido el ADN que adquirió la función de almacén de información genética y no su hermano, el ARN, se puede deber al hecho que es más flexible, al ser capaz de acomodar mutaciones genéticas sin desagregarse[60]. Hubo dudas sobre la estabilidad del ADN en condiciones como las de la tierra primordial, con una alta incidencia de rayos ultravioleta que pueden causar avalanchas de mutaciones genéticas, pero se determinó que en agua, el ADN es bastante estable[61].
Es posible que el ADN no se originó como un almacén de información genética, sino como un acumulador de fósforo, que es un elemento bastante raro en la naturaleza, pero crítico para los organismos ya que los compuestos que les proporcionan la energía contienen fósforo. Se ha determinado que una arquea, Haloferax volcanii, utiliza ADN duplicado para acumular fósforo para utilizarlo en tiempos de escasez del elemento, y es posible que el ADN apareció originalmente con este fin, y que sólo después se convirtió en portador de información genética[62]. La razón que el ADN sustituyó al ARN como portador de la información genética es que es más estable que el ARN y, por tanto, más idóneo para almacenar información[63].
Existen dudas acerca la edad relativa del ADN y el ARN: muchos consideran que el ARN apareció primero, y el ADN después, formado a partir del ARN; pero la fase intermedia entre ADN y ARN, lo que se llaman quimeras, biomoléculas compuestas por una unión de ADN y ARN, no son estables. Por tanto, es posible que el ADN y el ARN hayan aparecido más o menos contemporáneamente, de manera independiente[64].
Todavía estamos en el proceso de aprender cómo se pueden haber formado las primeras moléculas de ADN. En la naturaleza es muy poco probable que se forme una molécula de ADN (o ARN) desde cero. Es preciso entender cuáles piezas pueden haber formado estas moléculas, y cómo se pueden haber organizado en las cadenas en forma de hélice, típicas del ADN y ARN.
Primeros replicantes
En los organismos actuales, la formación del ADN y el ARN es facilitada por la presencia de un ejército de proteínas. Pero en los albores de la vida, antes de que existieran estas proteínas, ¿cómo pueden haberse formado las primeras moléculas de ARN? (El ADN probablemente vino después del ARN.)
Para obtener moléculas de ADN o ARN es necesario juntar de manera específica a una multitud de compuestos. Juntar ribosas y nucleobases para formar los nucleótidos (los “eslabones” de las cadenas) es difícil en condiciones prebióticas, pero no imposible. Existe por lo menos una ruta para lograrlo, en condiciones que pueden haber existido en la tierra primordial[65]. Fragmentos de ARN pueden formarse en la superficie de ciertas arcillas (ver arriba), y también en agua, bajo ciertas condiciones[66]. Los fragmentos de ARN, una vez formados, pueden juntarse para formar cadenas más largas en ambientes acuáticos ácidos, a temperaturas de menos de 70°C, sin la necesidad de la presencia de enzimas[67].
La probabilidad de que se forme el ARN o ADN en condiciones prebióticas es baja. Pero es posible que hubo ayuda. Por ejemplo, se determinó que en los tiempos que apareció la vida terrestre, las aguas superficiales de nuestro planeta contenían una alta concentración de sulfitos; estos son capaces de facilitar la formación del ARN[68]. También es posible que hubo cierto grado de autoorganización que ayudara: se observó que cortos trozos de polímeros con una estructura en forma de hélice, que tienen la capacidad de formar cristales líquidos (que tienen propiedades en parte de sólidos, en parte de líquidos), se alinean espontáneamente. Al alinearse, forman filamentos largos que posiblemente pudieron haber servido como base para la formación de las moléculas de ADN o ARN[69]. Para que se junten filamentos cortos para formar polímeros largos, es importante la presencia de moléculas que “peguen” un filamento a otro, tales como el etidio; en la ausencia de estas moléculas, los filamentos formarían anillos en lugar de filamentos rectos capaces de juntarse uno con otro[70].
Probablemente el ADN y el ARN, tal como los conocemos ahora, tuvieron originalmente una forma más sencilla[71]. Por ejemplo, los primeros replicantes (precursores del ADN y ARN) no necesariamente tenían las cuatro nucleobases que el ADN y el ARN tienen ahora. Es posible que originalmente hubo sólo dos. Esto simplificaría considerablemente el ensamblaje de los replicantes, especialmente considerando que una de las bases, la citosina (la letra C), se descompone fácilmente tan pronto se eleva la temperatura[72].
Entre los precursores del ARN (y el ADN), también pueden haber figurado polímeros distintos a éstos, pero capaces de almacenar información genética. Entre los posibles polímeros distintos se encuentran la triaminopirimidina[73], los APN (ácidos nucleicos basados en péptidos[74]), y una biomolécula denominada ATN[75]. Un péptido llamado beta-amiloide puede haber sido capaz de replicarse y transmitir información al descomponerse en los compuestos de los que se formó antes de replicarse[76]. También es posible que hayan existido otras nucleobases, por ejemplo una que se llama la triazina[77].
También se ha propuesto que los primeros biopolímeros no se formaron a partir de aminoácidos, ya que esto en la práctica es difícil en ausencia de enzimas, sino a partir de ácidos alfa hidróxidos[78].
Más exótica es la hipótesis que postula que los primeros replicantes pueden haber sido minerales, específicamente arcillas. En su estructura, un cristal de arcilla puede contener información en la forma de defectos cristalinos, o elementos varios incluidos en la estructura, etc.; esta información pudiera ser transmitida a nuevos cristales de arcilla, en un proceso que puede seleccionar las estructuras mejor adaptadas. Poco a poco, estos replicantes minerales pudieran haber incorporado moléculas biológicas, dando así origen al primer ARN[79]. Por poco probable que pueda parecer esta hipótesis, nos recuerda que en los albores de la vida existía sin duda una relación estrecha entre los minerales y los compuestos prebióticos.
Probablemente los primeros replicadores vinieron en una variedad de formas, de las que el futuro ARN era sólo una. Desconocemos cómo fue la posible rivalidad entre el ARN y sus competidores, y cuál fue la causa de que ganara el ARN[80]. Lo que sí se pudo determinar es que las proto-células contentivas de replicadores tenían mejor probabilidad de sobrevivir en el proceso de la selección natural que proto-células sin replicadores[81].
No está muy claro cómo pudieron replicarse los primeros replicadores. ¿Formaron cadenas moleculares complementarias, así como hace el ADN actual cuando se replica? Sin el apoyo de enzimas, esto es difícil[82]. Si se pudo formar una cadena complementaria, debe haber habido algún mecanismo para evitar que la cadena complementaria se quedara “pegada” a la cadena original[83]. Puede haber contribuido a solucionar esto, alguna ribozima capaz de elaborar ARN de una quiralidad opuesta[84] (o sea, distinto a su imagen especular, como nuestra mano izquierda es distinta a la derecha[85]), antes de que los organismos vivos empezaran a favorecer la quiralidad S [86]. También pueden haber ayudado cambios en temperatura: la cadena complementaria se formaría en aguas más frías, y se desprendería de la cadena original en aguas más cálidas[87]; de tales ambientes cambiantes volveremos a hablar en la próxima entrega.
¿O se construían cadenas nuevas a partir de la información genética codificada en los replicadores? Esto requiere de un mecanismo para traducir en aminoácidos la información genética codificada, mediante codones en el replicador; este mecanismo probablemente se pudo establecer gracias a una afinidad química entre el codón y el aminoácido correspondiente[88]. Para leer el código del ARN primordial y copiarlo, debe haber existido una manera de abrir la estructura helicoidal del ARN por suficiente tiempo. Sin el apoyo de las biomoléculas apropiadas es difícil abrir la estructura helicoidal, para que puedan entrar otras moléculas para leer el código del ARN y copiarlo; y además, en agua el ARN se mantiene abierto durante muy poco tiempo, insuficiente para el copiado. Pero se determinó que, en la presencia de solventes (por ejemplo, glicerol) que hacen más denso el medio acuático, el ARN se mantiene abierto más tiempo de manera que puede ser leído[89].
Se supone que al inicio ocurrían muchos errores de descodificación, aunque es posible que hubo mecanismos para minimizar estos errores[90]. En general, deben haberse dado inicialmente muchos errores de copiado en el proceso de replicación, ya que los mecanismos de replicación primordiales eran simples. Sin embargo, se determinó que en un mundo ARN (ver abajo) la fidelidad de copiado de las ribozimas pudo haber sido suficiente para que la replicación fuese efectiva. Además, se piensa que los primeros replicadores eran cortos, de manera que la cantidad de errores se pudo mantener baja[91]. También se determinó que, en poblaciones pequeñas, los errores de copiado pueden más bien ser beneficiosos para la evolución, al contrarrestar la deriva genética[92].
Todo lo arriba mencionado se basa en la suposición de que la vida se originó en un ambiente acuático, y que todas las reacciones biológicas ocurren en agua. Sin embargo, el agua no sólo hace posible la construcción de las biomoléculas: también es corrosiva, y puede causar, con el tiempo, la disolución de los enlaces químicos que mantienen unidas las piezas que componen las biomoléculas. Por tanto, se ha propuesto que la vida pudiera haberse desarrollado en un ambiente que no fuera acuoso: bien sea en otro planeta donde no hay agua, bien sea en la tierra, en un líquido distinto al agua, por ejemplo la formamida, que posiblemente se formó en la tierra primordial bajo la influencia de un bombardeo por partículas radioactivas[93]. Esta hipótesis, sin embargo, no explica cómo la vida, si de verdad se formó en tal líquido, hiciera el salto a un ambiente acuático, supuestamente corrosivo.
El mundo ARN
En los organismos actuales, para que haya un portador de la información genética y para que ésta pueda ser pasada a la siguiente generación, el ADN es fundamental, pero de por si insuficiente: la vida requiere no sólo de replicantes, sino también de catalizadores enzimáticos para controlar el metabolismo, o sea, la conversión de alimentos en energía para que se puedan desarrollar los procesos celulares, y en materia prima para la construcción de la célula. Por tanto, el descubrimiento, en los años ’60, de que el ARN, componente clave en la transferencia de información del ADN a las proteínas, también puede ser una enzima, abrió nuevos horizontes en la búsqueda del origen de la vida[94].
Así nació el concepto del “Mundo ARN” [95], según el cual la vida comenzó cuando la mezcla primordial dio origen a las ribozimas (moléculas de ARN que actúan como enzimas), que pueden a la vez replicarse y controlar el metabolismo. Un organismo autónomo en el Mundo ARN sólo es posible si el ARN pudiera asumir varias de las funciones actualmente desempeñadas por las proteínas, por ejemplo, la elaboración del ARN mismo. El descubrimiento imprevisto de las ribozimas, que catalizan reacciones en las células, similares a las enzimas, marcó el comienzo del interés actual en un mundo biológico sin proteínas. Hoy día, las ribozimas tienen poco uso, pero son vestigios de épocas remotas, cuando – antes de que existiera el ADN – el ARN estaba al mando de la actividad celular. Se demostró que las ribozimas pueden catalizar un número significativo de reacciones químicas diversas. Esto resultó en un mayor interés en la hipótesis de un Mundo ARN anterior al mundo actual de ADN/ARN/proteína. Cuando se determinó la estructura del ribosoma y se demostró que este es una ribozima (o sea, que puede actuar como una enzima)[96], quedó prácticamente confirmado el caso del Mundo ARN[97]. Ribozimas son capaces de replicarse y de mantener un sistema químico capaz de evolucionar, cumpliendo así con la definición de la vida mencionada al inicio de esta entrega[98]. En el laboratorio se logró obtener una ribozima que, en efecto, es capaz de sintetizar secuencias de ARN nuevas así como crear ARN nuevo mediante replicación[99].
En efecto, el ribosoma parece haber sido una pieza clave en el desarrollo de la vida: contiene vestigios de proto-genes que pueden haber codificado información necesaria para codificar y realizar su propia construcción y la de proteínas. De ser así, el ribosoma fue la biomolécula que permitió el desarrollo de la primera vida, antes de que apareciera el ADN[100].
Una característica de los biopolímeros dentro de las células es que están plegados, formando bolas. Esto aplica también al ARN y las ribozimas. Por ende, para que una ribozima pueda copiarse al ARN (y a si misma), debe poder abrir esta bola del ARN lo suficiente para leer el código genético. Se determinó que, en el Mundo ARN, una versión del ARN, capaz de leer tripletes de codones en lugar de los codones (las letras A, C, G y U) uno a la vez, debería haber sido capaz de leer moléculas de ARN plegadas[101].
Aun en nuestros días hay bacterias en las que ciertos tipos de ARN realizan actividades propias de las proteínas, lo que sugiere que cuando apareció la vida la presencia de proteínas no fue fundamental[102]. Sin embargo, proteínas primordiales (o más bien sus hermanos menores, los péptidos) pueden haber ayudado en los albores de la vida: el desarrollo del primer ARN puede haber ido de la mano de la aparición de otras biomoléculas, que pueden haber interactuado con el ARN y ayudado así en el desarrollo de los primeros organismos. Estas biomoléculas pueden haber sido unas proteínas sencillas[103], o enzimas primordiales[104]. Este concepto se llama el “Mundo de ARN/péptidos” [105]. Según esta hipótesis, sin los péptidos el primer ARN posiblemente ni hubiera podido formarse[106].
Para que los primeros organismos en el Mundo ARN hayan podido obtener la energía requerida, se ha postulado un mecanismo llamado termosíntesis, que es más sencillo que los mecanismos utilizados hoy en día (que incluyen, entre otros, la fotosíntesis) y que además dependería de una sola proteína simple, hipotética. Esta proteína, al estar expuesta a cambios de temperatura (por ejemplo en células de convección en el mar, cerca de fuentes calientes), pudiera mediar en la conversión de la molécula ADP en ATP, recargando así la “batería” del organismo[107].
Hacia el mundo ADN
Puesto que actualmente vivimos en un mundo “ADN/ARN/proteínas”, en algún momento debe haber ocurrido una evolución a este mundo a partir del Mundo ARN[108]. En un primer tiempo, el ARN empezó a producir proteínas que desempeñaron funciones que antes las desempeñaba el ARN (vestigios de este paso aun hoy día se pueden observar en ciertos hongos[109]). Después debe haber aparecido el ADN. No sabemos cómo fue esta transición, pero el ribosoma jugó probablemente un papel clave[110]. Por ejemplo, se demostró que es posible que una ribozima (una molécula de ARN que actúa como enzima) se puede convertir en una deoxiribozima, o sea, una enzima que actúa en el mundo ADN, transmitiendo en la conversión la información genética del Mundo ARN al Mundo ADN[111].
Todo esto debe haber ocurrido temprano, antes de que viviera el LUCA, el último ancestro común de todas las formas de vida que en la actualidad habitan la tierra (en la entrega anterior vimos que el LUCA debe haber vivido hace unos 3500-3800 millones de años).
Conclusión
La química de los procesos biológicos es compleja, pero debe haber tenido unos inicios sencillos. Con el desarrollo del concepto del Mundo ARN se ha hecho un gran avance hacia el entendimiento de cómo se formaron las primeras moléculas de la vida.
Al inicio de esta entrega vimos que hay cuatro características con las que tiene que contar un organismo para que pueda ser considerado vivo: debe tener un envoltorio, ser capaz de dividirse, contar con material genético, y tener un mecanismo para generar energía. Con el concepto del Mundo ARN y lo que sabemos de las membranas lipídicas se tiene cierto entendimiento – todavía incompleto – de las primeras tres características. Nos falta considerar la energía de los organismos, o sea, su metabolismo. De esto hablaremos en la próxima entrada de este blog.
Nota: la imagen en el encabezado de esta entrega muestra una proto-célula, mostrando la membrana y, en el interior, el material genético (ARN). Las manchas blanco-azules afuera y dentro de la proto-célula son nucleótidos. El diámetro de la proto-célula es de unos 100 nanómetros (un milímetro equivale a un millón de nanómetros). Crédito: Janet Iwasa, Szostak Group, MGH/Harvard. Fuentes: https://www.nsf.gov/news/mmg/mmg_disp.jsp?med_id=65925, y http://www.macmillanhighered.com/BrainHoney/Resource/6716/digital_first_content/trunk/test/hillis2e/asset/img_ch4/c04_fig19.html.
[1] Singer, E., 2015. How structure arose in the primordial soup. Quanta Magazine. www.quantamagazine.org/20150416howstructureevolvedintheprimordialsoup.
[2] Rasmussen, S., Chen, L., Deamer, D., Krakauer, D.C., Packard, N.H., Stadler, P.F. y Bedau, M.A., 2004. Transitions from nonliving to living matter. Science, 303 (5660), 963-965. http://science.sciencemag.org/content/303/5660/963. Szathmary, E., 2005. Life: In search of the simplest cell. Nature, 433, 469-470. www.nature.com/articles/433469a.
[3] Ver: https://es.wikipedia.org/wiki/Tit%C3%A1n_(sat%C3%A9lite).
[4] Martin, W. y Russell, M.J., 2003. On the origins of cells: a hypothesis for the evolutionary transitions from abiotic geochemistry to chemoautotrophic prokaryotes, and from prokaryotes to nucleated cells. Philosophical Transactions of the Royal Society of London, B, 358, 59-85. http://rstb.royalsocietypublishing.org/content/358/1429/59. Caforio, A. y otros, 2018. Converting Escherichia coli into an archaebacterium with a hybrid heterochiral membrane. Proceedings of the National Academy of Sciences, 115 (14), 3704-3709. www.pnas.org/content/115/14/3704.
[5] Ver: https://es.wikipedia.org/wiki/Fosfol%C3%ADpido.
[6] Hargreaves, W.R., Mulvihill, S.J. y Deamer, D.W., 1977. Synthesis of phospholipids and membranes in prebiotic conditions. Nature, 266, 78-80. www.nature.com/articles/266078a0.
[7] Russell, M.J., 2003. The importance of bein-g alkaline. Science, 302 (5645), 580-581. http://science.sciencemag.org/content/302/5645/580.
[8] Mansy, S.S., Schrum, J.P., Krishnamurthy, M., Tobé, S., Treco, D.A. y Szostak, J.W., 2008. Template-directed synthesis of a genetic polymer in a model protocell. Nature, 454, 122-125. www.nature.com/articles/nature07018.
[9] Chen, I.A., Roberts, R.W. y Szostak, J.W., 2004. The emergence of competition between model protocells. Science, 305, 1474-1476. http://science.sciencemag.org/content/305/5689/1474. Hentrich, C. y Szostak, J.W., 2014. Controlled growth of filamentous fatty acid vesicles under flow. Langmuir, 30 (49), 14916-14925. https://pubs.acs.org/doi/10.1021/la503933x.
[10] Chen, I.A. y Szostak, J.W., 2004. A kinetic study of the growth of fatty acid vesicles. Biophysical Journal, 87, 988-998. www.sciencedirect.com/science/article/pii/S0006349504735821. Kamimura, A. y Kaneko, K., 2010. Reproduction of a protocell by replication of a minority molecule in a catalytic reaction network. Physical Review Letters, 105, 268103. https://journals.aps.org/prl/abstract/10.1103/PhysRevLett.105.268103.
[11] Giomi, L. y DeSimone, A., 2014. Spontaneous division and motility in active nematic droplets. Physical Review Letters, 112, 147802. https://journals.aps.org/prl/abstract/10.1103/PhysRevLett.112.147802.
[12] Hanczyc, M.M., Fujikawa, S.M. y Szostak, J.W., 2003. Experimental models of primitive cellular compartments: encapsulation, growth, and division. Science, 302, 618-622. http://science.sciencemag.org/content/302/5645/618.
[13] Leaver, M., Domínguez-Cuevas, P., Coxhead, J.M., Daniel, R.A. y Errington, J., 2009. Life without a wall or division machine in Bacillus subtilis. Nature, 457 (7231), 849-853. www.nature.com/articles/nature07742. Mercier, R., Kawai, Y. y Errington, J., 2013. Excess membrane synthesis drives a primitive mode of cell proliferation. Cell, 152 (5), 942-944. www.sciencedirect.com/science/article/pii/S0092867413001359.
[14] Stano, P., D’Aguanno, E., Bolz, J., Fahr, A. y Luisi, P.L., 2013. A remarkable self‐organization process as the origin of primitive functional cells. Angewandte Chemie, 52 (50), 13397-13400. https://onlinelibrary.wiley.com/doi/abs/10.1002/anie.201306613.
[15] García-Ruiz, J.M., Nakouzi,E., Kotopoulou, E., Tamborrino, L. y Steinbock, O., 2017. Biomimetic mineral self-organization from silica-rich spring waters. Science Advances, 3, e1602285. http://advances.sciencemag.org/content/3/3/e1602285.
[16] Strulson, C.A., Molden, R.C., Keating, C.D. y Bevilacqua, P.C., 2012. RNA catalysis through compartmentalization. Nature Chemistry, 4, 941-946. www.nature.com/articles/nchem.1466.
[17] Martin, W. y Russell, M.J., 2003. On the origins of cells: a hypothesis for the evolutionary transitions from abiotic geochemistry to chemoautotrophic prokaryotes, and from prokaryotes to nucleated cells. Philosophical Transactions of the Royal Society of London, B, 358, 59-85. http://rstb.royalsocietypublishing.org/content/358/1429/59.
[18] Mulkidjanian, A.Y., 2009. On the origin of life in the Zinc world: 1. Photosynthesizing, porous edifices built of hydrothermally precipitated zinc sulfide as cradles of life on Earth. Biology Direct, 4, 26. https://biologydirect.biomedcentral.com/articles/10.1186/1745-6150-4-26. Mulkidjanian, A.Y. y Galperin, M.Y., 2009. On the origin of life in the Zinc world. 2. Validation of the hypothesis on the photosynthesizing zinc sulfide edifices as cradles of life on Earth. Biology Direct, 4, 27. https://biologydirect.biomedcentral.com/articles/10.1186/1745-6150-4-27.
[19] Watson, J.D. y Crick, F.H.C., 1953. Molecular structure of nucleic acids. Nature, 171 (4356), 737-738. www.nature.com/articles/171737a0.pdf.
[20] Ver: https://es.wikipedia.org/wiki/%C3%81cido_desoxirribonucleico.
[21] Ver: https://es.wikipedia.org/wiki/%C3%81cido_ribonucleico.
[22] Ver: https://es.wikipedia.org/wiki/Transcripci%C3%B3n_gen%C3%A9tica.
[23] Ver: https://es.wikipedia.org/wiki/Traducci%C3%B3n_(gen%C3%A9tica).
[24] Petrov, A.S. y otros, 2015. History of the ribosome and the origin of translation. Proceedings of the National Academy of Sciences, 112 (50), 15396-15401. www.pnas.org/content/pnas/112/50/15396.full.pdf.
[25] Johnson, D.B.F. y Wang, L., 2010. Imprints of the genetic code in the ribosome. Proceedings of the National Academy of Sciences, 107 (18), 8298-8303. www.pnas.org/content/pnas/107/18/8298.full.pdf.
[26] Ver: https://es.wikipedia.org/wiki/Base_nitrogenada.
[27] Stairs, S., Nikmal, A., Bučar, D.-K., Zheng, S.-L., Szostak, J.W. y Powner, M.W., 2017. Divergent prebiotic synthesis of pyrimidine and 8-oxo-purine ribonucleotides. Nature Communications, 8, 15270. www.nature.com/articles/ncomms15270. Becker, S., Thoma, I., Deutsch, A., Gehrke, T., Mayer, P., Zipse, H. y Carell, T., 2016. A high-yielding, strictly regioselective prebiotic purine nucleoside formation pathway. Science, 352 (6287), 833-836. http://science.sciencemag.org/content/352/6287/833.
[28] Ricardo, A., Carrigan, M.A., Olcott, A.N. y Benner, S.A., 2004. Borate minerals stabilize ribose. Science, 303, 196. http://science.sciencemag.org/content/303/5655/196.
[29] Ricardo, A. y Szostak, J.W., 2009. The Origin of Life on Earth. Scientific American, septiembre 2009, 54-61. www.scientificamerican.com/article/origin-of-life-on-earth.
[30] Patel, B.H., Percivalle, C., Ritson, D.J., Duffy, C.D. y Sutherland, J.D., 2015. Common origins of RNA, protein and lipid precursors in a cyanosulfidic protometabolism. Nature Chemistry, 7, 301-307. www.nature.com/articles/nchem.2202.
[31] Ver: https://es.wikipedia.org/wiki/P%C3%A9ptido; https://es.wikipedia.org/wiki/Prote%C3%ADna.
[32] Leman, L., Orgel, L. y Ghadiri, M.R., 2004. Carbonyl sulphide-mediated prebiotic formation of peptides. Science, 306, 283-286. http://science.sciencemag.org/content/306/5694/283.full. Huber, C., Eisenreich, W., Hecht, S. y Wächtershäuser, G., 2003. A possible primordial peptide cycle. Science, 301, 938-940. http://science.sciencemag.org/content/301/5635/938.full. Huber, C. y Wächtershäuser, G., 1998. Peptides by activation of amino acids with CO on (Ni,Fe)S surfaces: implications for the origin of life. Science, 281, 670-672. http://science.sciencemag.org/content/281/5377/670.full.
[33] Kim, J.D., Pike, D.H., Tyryshkin A.M., Swapna, G.V.T., Raanan, H., Montelione‡, G.T., Nanda, V. y Falkowski, P.G., 2018. Minimal heterochiral de novo designed 4Fe–4S binding peptide capable of robust electron transfer. Journal of the American Chemical Society, gky600. https://academic.oup.com/nar/advance-article/doi/10.1093/nar/gky600/5054862.
[34] Carter, C.W. y Wills, P.R., 2018. Hierarchical groove discrimination by Class I and II aminoacyl-tRNA synthetases reveals a palimpsest of the operational RNA code in the tRNA acceptor-stem bases. Nucleic Acids Research, 140 (36), 11210-11213. https://pubs.acs.org/doi/10.1021/jacs.8b07553#.
[35] Gibard, C., Bhowmik, S., Karki, M., Kim, E.-K. y Krishnamurthy, R., 2017. Phosphorylation, oligomerization and self-assembly in water under potential prebiotic conditions. Nature Chemistry, 10, 212-217. www.nature.com/articles/nchem.2878.
[36] Longo, L.M., Lee, J. y Blaber, M., 2013. Simplified protein design biased for prebiotic amino acids yields a foldable, halophilic protein. Proceedings of the National Academy of Sciences, 110 (6), 2135-2139. www.pnas.org/content/110/6/2135.
[37] Mamajanov, I. y Cody, G.D., 2017. Protoenzymes: the case of hyperbranched polyesters. Philosophical Transactions of the Royal Society A, 375, 20160357. http://rsta.royalsocietypublishing.org/content/375/2109/20160357.
[38] Meringer, M. y Cleaves, H.J., 2017. Exploring astrobiology using in silico molecular structure generation. Philosophical Transactions of the Royal Society A, 375, 20160344. http://rsta.royalsocietypublishing.org/content/375/2109/20160344.
[39] Lancet, D., Zidovetzki, R. y Markovitch, O., 2018. Systems protobiology: origin of life in lipid catalytic networks. Journal Royal Society Interface, 15, 20180159. http://rsif.royalsocietypublishing.org/content/royinterface/15/144/20180159.full.pdf.
[40] Hazen, R.M., 2013. Paleomineralogy of the Hadean Eon: A preliminary species list. American Journal of Science, 313, 807-843. www.ajsonline.org/content/313/9/807.short.
[41] Ferris, J.P., 2005. Mineral catalysis and prebiotic synthesis: montmorillonite-catalyzed formation of RNA. Elements, 1 (3), 145-149. https://pubs.geoscienceworld.org/msa/elements/article-abstract/1/3/145/137565/mineral-catalysis-and-prebiotic-synthesis.
[42] Hazen, R.M., 2005. Genesis: rocks, minerals, and the geochemical origin of life. Elements, 1 (3), 135-137. https://pubs.geoscienceworld.org/msa/elements/article-abstract/1/3/135/137558/genesis-rocks-minerals-and-the-geochemical-origin.
[43] Sowerby, S.J., Edelwirth, M. Heckl, W.M., 1998. Self-assembly at the prebiotic solid−liquid interface: structures of self-assembled monolayers of adenine and guanine bases formed on inorganic surfaces. Journal of Physical Chemistry B, 102 (30), 5914-5922. https://pubs.acs.org/doi/abs/10.1021/jp980684l?journalCode=jpcbfk#.
[44] Sommer, A.P., Zhu, D. y Fecht, H.-J., 2008. Genesis on diamonds. Crystal Growth & Design, 8 (8), 2628-2629. https://pubs.acs.org/doi/pdfplus/10.1021/cg8005037. Sommer, A.P., Jaganathan, S. , Maduro, M.R., Hancke, K., Janni, W. y Fecht, H.-J., 2016. Genesis on diamonds II: contact with diamond enhances human sperm performance by 300%. Annals of Translational Medicine, ;4 (20), 407. http://atm.amegroups.com/article/view/11322/pdf.
[45] Hansma, H.G., 2007. Mica and the origin of life: cells without membranes. American Society for Cell Biology, 47th Annual Meeting, abstract 1910. www.ascb.org/wp-content/uploads/2015/12/2007-abstracts.pdf. Hansma, H.G., 2010. Possible origin of life between mica sheets. Journal of Theoretical Biology, 266 (1), 175-188. www.sciencedirect.com/science/article/pii/S0022519310003061.
[46] Hazen, 2005. Ver nota 42.
[47] Brasier, M.D., Matthewman, R., McMahon, S. y Wacey, D., 2011. Pumice as a remarkable substrate for the origin of life. Astrobiology, 11 (7). www.liebertpub.com/doi/pdf/10.1089/ast.2010.0546.
[48] Sleep, N.H., Bird, D.K. y Pope, E.C., 2011. Serpentinite and the dawn of life. Philosophical Transactions f the Royal Society B, 366, 2857-2869. http://rstb.royalsocietypublishing.org/content/royptb/366/1580/2857.full.pdf.
[49] Pontefract, A., Osinski, G.R., Cockell, C.S., Moore, C.A., Moores, J.E. y Southam, G., 2014. Impact-generated endolithic habitat within crystalline rocks of the Haughton impact structure, Devon island, Canada. Astrobiology, 14 (6), 522-533. www.liebertpub.com/doi/10.1089/ast.2013.1100.
[50] Bryce, C., Horneck, G., Rabbow, E., Edwards, H. y Cockell, C., 2015. Impact shocked rocks as protective habitats on an anoxic early Earth. International Journal of Astrobiology, 14 (1), 115-122. www.cambridge.org/core/journals/international-journal-of-astrobiology/article/impact-shocked-rocks-as-protective-habitats-on-an-anoxic-early-earth/AF74F27D4D1E0B5970BA16C357069C81.
[51] Schmieder, M. y Jourdan, F., 2013. The Lappajärvi impact structure (Finland): Age, duration of crater cooling, and implications for early life. Geochimica et Cosmochimica Acta, 112, 321-339. www.sciencedirect.com/science/article/pii/S0016703713001105. Williams, F.A., Kelley, S.P., Gilmour, I., Jolley, D.W. y Gilmour, M., 2013. The Boltysh impact crater, Ukraine: smectites from the crater-fill suevites. EPSC Abstracts, 8, EPSC2013-305. http://meetingorganizer.copernicus.org/EPSC2013/EPSC2013-305.pdf.
[52] Sapers, H.M., Osinski, G.R., Banerjee, N.R. y Preston, L.J., 2014. Enigmatic tubular features in impact glass. Geology, 42 (6), 471-474. https://pubs.geoscienceworld.org/gsa/geology/article-abstract/42/6/471/131521/enigmatic-tubular-features-in-impact-glass.
[53] Ball, R. y Brindley, J., 2017. Toy trains, loaded dice and the origin of life: dimerization on mineral surfaces under periodic drive with Gaussian inputs. Royal Society open science, 4, 170141. http://rsos.royalsocietypublishing.org/content/royopensci/4/11/170141.full.pdf.
[54] Chatterjee, S., 2013. Impact, RNA-protein world, and the endoprebiotic origin of life. Geological Society of America Abstracts with Programs, 45 (7), 694. https://gsa.confex.com/gsa/2013AM/webprogram/Paper222699.html. Chan, Q.H.S. y otros, 2018. Organic matter in extraterrestrial water-bearing salt crystals. Science Advances, 4, eaao3521. http://advances.sciencemag.org/content/advances/4/1/eaao3521.full.pdf.
[55] Smith, J.V., 2005. Geochemical influences on life’s origins and evolution. Elements, 1 (3), 151-156. https://pubs.geoscienceworld.org/msa/elements/article-abstract/1/3/151/137568/geochemical-influences-on-life-s-origins-and.
[56] Hanczyc y otros, 2003. Ver nota 12.
[57] Mulkidjanian, A.Y., Bychkov, A.Y., Dibrova, D.V., Galperin, M.Y. y Koonin, E.V., 2012. Origin of first cells at terrestrial, anoxic geothermal fields. Proceedings of the National Academy of Sciences, 109 (14), E821-E830. http://www.pnas.org/content/109/14/E821.
[58] Ver: https://es.wikipedia.org/wiki/Aleksandr_Oparin.
[59] Miller, S.L., 1953. A production of amino acids under possible primitive Earth conditions. Science, 117 (3046), 528-529. http://science.sciencemag.org/content/117/3046/528.
[60] Zhou, H. y otros, 2016. m1A and m1G disrupt A-RNA structure through the intrinsic instability of Hoogsteen base pairs. Nature Structural & Molecular Biology, 23, 803-810. www.nature.com/articles/nsmb.3270.
[61] He, Y., Wu, C. y Kong, W., 2004. Photophysics of methyl-substituted uracils and thymines and their water complexes in the gas phase. Journal of Physical Chemistry A, 108 (6), 943-949. https://pubs.acs.org/doi/abs/10.1021/jp036553o.
[62] Zerulla, K., Chimileski, S., Näther, D., Gophna, U., Papke, R.T. y Soppa, J., 2014. DNA as a phosphate storage polymer and the alternative advantages of polyploidy for growth or survival. PLoS ONE, 9 (4), e94819. https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0094819.
[63] Jogalekar, A., 2014. How a college student can derive the RNA world hypothesis from scratch. https://blogs.scientificamerican.com/the-curious-wavefunction/how-a-college-student-can-derive-the-rna-world-hypothesis-from-scratch.
[64] Gavette, J.V., Stoop, M., Hud, N.V. y Krishnamurthy, R., 2016. RNA-DNA chimeras in the context of an RNA world transition to an RNA/DNA world. Angewandte Chemie International Edition, 55 (42), 13204-13209. https://onlinelibrary.wiley.com/doi/full/10.1002/anie.201607919.
[65] Powner, M.W., Gerland, B. y Sutherland, J.D., 2009. Synthesis of activated pyrimidine ribonucleotides in prebiotically plausible conditions. Nature, 459, 239–242. www.nature.com/articles/nature08013.
[66] Ricardo y Szostak, 2009. Ver nota 29.
[67] Pino, S., Ciciriello, F., Costanzo, G. y Di Mauro, E., 2008. Nonenzymatic RNA ligation in water. The Journal of Biological Chemistry, 283 (52), 36494-36503. www.jbc.org/content/283/52/36494.full.pdf.
[68] Ranjan, S., Todd, Z.R., Sutherland, J.D. y Sasselov, D.D., 2018. Sulfidic anion concentrations on early Earth for surficial origins-of-life chemistry. Astrobiology, 18, 8. www.liebertpub.com/doi/pdf/10.1089/ast.2017.1770.
[69] Nakata, M., Zanchetta, G., Chapman, B.D., Jones, C.D., Cross, J.O., Pindak, R., Bellini, T. y Clark, N.A., 2007. End-to-end stacking and liquid crystal condensation of 6– to 20–base pair DNA duplexes. Science, 318, 1276-1279. http://science.sciencemag.org/content/318/5854/1276.
[70] Horowitz, E.D., Engelhart, A.E., Chen, M.C., Quarles, K.A., Smith, M.W., Lynn, D.G. y Hud, N.V., 2010. Intercalation as a means to suppress cyclization and promote polymerization of base-pairing oligonucleotides in a prebiotic world. Proceedings of the National Academy of Sciences, 107 (12), 5288-5293. www.pnas.org/content/107/12/5288.
[71] Hud, N.V., Cafferty, B.J., Krishnamurthy, R. y Williams, L.D., 2013. The origin of RNA and “my grandfather’s axe”. Chemistry & Biology, 20 (4), 466-474. www.sciencedirect.com/science/article/pii/S1074552113001154.
[72] Reader, J.S. y Joyce, G.F., 2002. A ribozyme composed of only two different nucleotides. Nature, 420, 841-844. www.nature.com/articles/nature01185.
[73] Cafferty, B.J., Gallego, I., Chen, M.C., Farley, K.I., Eritja, R., y Hud, N.V., 2013. Efficient self-assembly in water of long noncovalent polymers by nucleobase analogues. Journal of the American Chemical Society, 135, 2447-2450. https://pubs.acs.org/doi/10.1021/ja312155v. Chen, M.C., Cafferty, B.J., Mamajanov, I., Gallego, I., Khanam, J., Krishnamurthy, R., y Hud, N.V., 2014. Spontaneous prebiotic formation of a beta-ribofuranoside that self-assembles with a complementary heterocycle. Journal of the American Chemical Society, 136, 5640-5646. https://pubs.acs.org/doi/10.1021/ja410124v.
[74] Plöger T.A. y von Kiedrowski, G., 2011. Improved large‐scale liquid‐phase synthesis and high‐temperature NMR characterization of short (F‐)PNAs. Helvetica Chimica Acta, 94 (11), 1952-1980. https://onlinelibrary.wiley.com/doi/abs/10.1002/hlca.201100243. Plöger T.A. y von Kiedrowski, G., 2014. A self-replicating peptide nucleic acid. Organic & Biomolecular Chemistry, 12, 6908-6914. https://pubs.rsc.org/en/Content/ArticleLanding/2014/OB/C4OB01168F#!divAbstract.
[75] Orgel, L.E., 2004. Prebiotic chemistry and the origin of the RNA world. Critical Reviews in Biochemistry and Molecular Biology, 39 (2), 99-123. www.tandfonline.com/doi/abs/10.1080/10409230490460765.
[76] Maury, C.P.J., 2015. Origin of life. Primordial genetics: Information transfer in a pre-RNA world based on self-replicating beta-sheet amyloid conformers. Journal of Theoretical Biology, 382, 292-297. www.sciencedirect.com/science/article/pii/S0022519315003379.
[77] Mittapalli, G.K., Reddy, K.R., Xiong, H., Munoz, O., Han, B., De Riccardis, F., Krishnamurthy, R. y Eschenmoser, A., 2007. Mapping the landscape of potentially primordial informational oligomers: oligodipeptides and oligodipeptoids tagged with triazines as recognition elements. Angewandte Chemie, 46 (14), 2470-2477. https://onlinelibrary.wiley.com/doi/abs/10.1002/anie.200603207.
[78] Chandru, K., Guttenberg, N., Giri, C., Hongo, Y., Butch, C., Mamajanov, I. y Cleaves, J., 2018. Simple prebiotic synthesis of high diversity dynamic combinatorial polyester libraries. Communications Chemistry, 1, 30. www.nature.com/articles/s42004-018-0031-1.
[79] Cairns-Smith, A.G., 2005. Sketches for a mineral genetic material. Elements, 1 (3), 157-161. https://pubs.geoscienceworld.org/msa/elements/article-abstract/1/3/157/137571.
[80] Cleaves, H.J., Meringer, M. y Goodwin, J., 2015. 227 Views of RNA: Is RNA unique in its chemical isomer space? Astrobiology, 15, 7, 538-558. http://online.liebertpub.com/doi/pdfplus/10.1089/ast.2014.1213.
[81] Bianconi, G., Zhao, K., Chen, I.A., y Nowak, M.A., 2013. Selection for replicases in protocells. PLoS Computational Biology, 9, e1003051. https://journals.plos.org/ploscompbiol/article?id=10.1371/journal.pcbi.1003051.
[82] Zhang, W., Tam, C.P., Wang, J. y Szostak, J.W., 2016. Unusual base-pairing interactions in monomer-template complexes. ACS Central Science, 2, 916-926. https://pubs.acs.org/doi/pdfplus/10.1021/acscentsci.6b00278.
[83] Taylor, W.R., 2005. Stirring the primordial soup. Nature, 434, 705. www.nature.com/articles/434705a.
[84] Sczepanski, J.T. y Joyce, GF., 2014. A cross-chiral RNA polymerase ribozyme. Nature, 515, 440-442. www.nature.com/articles/nature13900.
[85] Ver: https://es.wikipedia.org/wiki/Quiralidad_(qu%C3%ADmica).
[86] Everts, S., 2013. Mirror molecules. Scientific American, 308, 78-81. www.nature.com/scientificamerican/journal/v308/n5/full/scientificamerican0513-78.html. Dreiling, J.M. y Gay, T.J., 2014. Chirally sensitive electron-induced molecular breakup and the Vester-Ulbricht hypothesis. Physical Review Letters, 113, 118103. https://journals.aps.org/prl/abstract/10.1103/PhysRevLett.113.118103. Soai, K., Shibata, T., Morioka, H. y Choji, K., 1995. Asymmetric autocatalysis and amplification of enantiomeric excess of a chiral molecule. Nature, 378, 767-768. www.nature.com/articles/378767a0.
[87] Ricardo y Szostak, 2009. Ver nota 29.
[88] Polyansky, A.A., Hlevnjak, M. y Zagrovic, B., 2013. Proteome-wide analysis reveals clues of complementary interactions between mRNAs and their cognate proteins as the physicochemical foundation of the genetic code, RNA Biology, 10 (8), 1248-1254. www.tandfonline.com/doi/abs/10.4161/rna.25977. Hajnic, M., Iregui Osorio, J. y Zagrovic, B., 2014. Computational analysis of amino acids and their sidechain analogs in crowded solutions of RNA nucleobases with implications for the mRNA–protein complementarity hypothesis. Nucleic Acids Research, 42 (21), 12984-12994. https://academic.oup.com/nar/article/42/21/12984/2902673.
[89] He, C., Gállego, I., Laughlin, B., Grover, M.A. y Hud, N.V., 2017. A viscous solvent enables information transfer from genelength nucleic acids in a model prebiotic replication cycle. Nature Chemistry, 9, 318-324. www.nature.com/articles/nchem.2628.
[90] Lehmann, J., Cibils, M. y Libchaber, A., 2009. Emergence of a code in the polymerization of amino acids along RNA templates. PLoS ONE, 4 (6), e5773. https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0005773.
[91] Kun, A., Santos, M. y Szathmáry, E., 2005. Real ribozymes suggest a relaxed error threshold. Nature Genetics, 37,1008-1011. www.nature.com/articles/ng1621. Holmes, E.C., 2005. On being the right size. Nature Genetics, 37, 923-924. www.nature.com/articles/ng0905-923.
[92] Krakauer, D.C. y Sasaki, A., 2002. Noisy clues to the origin of life. Proceedings of the Royal Society B, 269, 2423-2428. http://tuvalu.santafe.edu/~krakauer/Site/Publications_files/Krakauer2002a.pdf.
[93] Adam, Z.R., Hongo, Y., Cleaves II, H.J., Yi, R., 3, Fahrenbach, A.C., Yoda, I. y Aono, M., 2018. Estimating the capacity for production of formamide by radioactive minerals on the prebiotic Earth. Scientific Reports, 8, 265. www.nature.com/articles/s41598-017-18483-8.pdf.
[94] Pressman, A., Blanco, C. y Cheri, I.A., 2015. The RNA world as a model system to study the origin of life. Current Biology, 25, R953–R963. www.sciencedirect.com/science/article/pii/S0960982215006818.
[95] Gilbert, W., 1986. Origin of life: the RNA world. Nature, 319, 618. www.nature.com/articles/319618a0. Ver también: www.the-scientist.com/research/scientists-debate-rnas-role-at-beginning-of-life-on-earth-57549.
[96] Steitz, T.A. y Moore, P.B., 2003. RNA, the first macromolecular catalyst: the ribosome is a ribozyme. Trends in Biochemical Sciences, Steitz, T.A., & Moore, P.B. (2003). RNA, the first macromolecular catalyst: the ribosome is a ribozyme. Trends in biochemical sciences, 28 (8), 411-418. www.cell.com/trends/biochemical-sciences/fulltext/S0968-0004(03)00169-5.
[97] Orgel 2004. Ver nota 75. Lee, E.R., Baker, J.L., Weinberg, Z., Sudarsan, N. y Breaker, R.R., 2010. An allosteric self-splicing ribozyme triggered by a bacterial second messenger. Science, 329 (5993), 845-848. http://science.sciencemag.org/content/329/5993/845.
[98] Lincoln, T.A. y Joyce, G.F., 2009. Self-sustained replication of an RNA enzyme. Science, 323, 1229-1232. http://science.sciencemag.org/content/sci/323/5918/1229.full.pdf. Vaidya, N., Manapat, M.L., Chen, I.A., Xulvi-Brunet, R., Hayden, E.J. y Lehman, N., 2012. Spontaneous network formation among cooperative RNA replicators. Nature, 491, 72-77. www.nature.com/articles/nature11549.
[99] Horning, D.P. y Joyce, G.F., 2016. Amplification of RNA by an RNA polymerase ribozyme. Proceedings of the National Academy of Sciences, 113 (35), 9786-9791. www.pnas.org/content/pnas/113/35/9786.full.pdf.
[100] Root-Bernstein, M. y Root-Bernstein, R., 2015. The ribosome as a missing link in the evolution of life. Journal of Theoretical Biology, 367, 130-158. www.sciencedirect.com/science/article/pii/S0022519314006778. Ver también: https://theconversation.com/never-mind-the-selfish-gene-ribosomes-are-the-missing-link-35409.
[101] Attwater, J., Raguram, A., Morgunov, A.S., Gianni, E. y Holliger, P., 2018. Ribozyme-catalysed RNA synthesis using triplet building blocks. eLife, 7, e35255. https://elifesciences.org/articles/35255.
[102] Sudarsan, N., Lee, E.R., Weinberg Z., Moy, R.H., Kim, J.N., Link, K.H. y Breaker, R.R., 2008. Riboswitches in eubacteria sense the second messenger cyclic di-GMP. Science, 321 (5887), 411-413. http://science.sciencemag.org/content/321/5887/411.
[103] Harish, A. y Caetano-Anollés, G., 2012. Ribosomal history reveals origins of modern protein synthesis. PLoS ONE, 7 (3), e32776. https://journals.plos.org/plosone/article/file?id=10.1371/journal.pone.0032776&type=printable.
[104] Li, L., Francklyn, C. y Carter, C.W., 2013. Aminoacylating urzymes challenge the RNA World hypothesis. The Journal of Biological Chemistry, 288, 26856-26863. www.jbc.org/content/288/37/26856.full.pdf. Carter, C.W. y Wills, P.R., 2018. Interdependence, reflexivity, fidelity, impedance matching, and the evolution of genetic coding. Molecular Biology and Evolution, 35 (2), 269-286. https://academic.oup.com/mbe/article/35/2/269/4430325.
[105] Taran, O. y otros, 2017. Expanding the informational chemistries of life: peptide/RNA networks. Philosophical Transactions of the Royal Society A, 375, 20160356. http://rsta.royalsocietypublishing.org/content/375/2109/20160356.
[106] Wills, P.R. y Carter, C.W., 2018. Insuperable problems of the genetic code initially emerging in an RNA world. BioSystems, 164, 155-166. www.sciencedirect.com/science/article/pii/S0303264717302332.
[107] Muller, A.W.J., 2005. Thermosynthesis as energy source for the RNA World: A model for the bioenergetics of the origin of life. BioSystems, 82, 93-102. www.sciencedirect.com/science/article/pii/S030326470500078X.
[108] Lilley, D.M.J. y Sutherland, J., 2011. The chemical origins of life and its early evolution: an introduction. Philosophical Transactions of the Royal Society of London, B, 366, 2853-2856. http://rstb.royalsocietypublishing.org/content/royptb/366/1580/2853.full.pdf.
[109] Paukstelis, P.J., Chen, J.-H., Chase, E., Lambowitz, A.M. y Golden, B.L., 2008. Structure of a tyrosyl-tRNA synthetase splicing factor bound to a group I intron RNA. Nature, 451, 94-97. www.nature.com/articles/nature06413.
[110] Meléndez-Hevia, E., 2009. From the RNA world to the DNA-protein world: clues to the origin and early evolution of life in the ribosome. Journal of Biosciences, 34, 825. https://rd.springer.com/article/10.1007%2Fs12038-009-0095-2.
[111] Paul, N., Springsteen, G. y Joyce, G.F., 2006. Conversion of a ribozyme to a deoxyribozyme through in vitro evolution. Chemistry & Biology, 13, 329-338. www.cell.com/action/showPdf?pii=S1074-5521%2806%2900042-1.

Un comentario en “Los albores de la vida (3): Amanecer”