
Los pinzones de las islas Galápagos alcanzaron fama mundial cuando el naturalista inglés Charles Darwin, quien los describió durante su expedición científica a bordo del barco Beagle, se dio cuenta que las distintas especies de pinzón que encontró en aquellas islas debían tener el mismo ancestro, al ser parecidas entre sí aunque cada especie contaba con su pico característico. Sin embargo, Darwin no pudo explicar, armado únicamente con su teoría de la selección natural[i], cómo aparecen especies nuevas a partir de especies anteriores. Sólo ahora somos capaces de explicar de manera bastante satisfactoria cómo ocurre esta especiación.
En el post anterior se habló del mecanismo de la selección natural, propuesto por Darwin para explicar cómo pueden cambiar en el tiempo los seres vivientes. Sin embargo, Darwin no explicó satisfactoriamente cómo pueden aparecer especies nuevas: el asunto es básicamente que cada especie está adaptada a su entorno, y si algún ejemplar de una especie nace con alguna mutación que le confiere una característica nueva, esto implica que no estaría adaptado de manera óptima a su entorno, y perdería en la lucha por la sobrevivencia. Obviamente, la sola acumulación de cambios mediante la selección natural no es suficiente para explicar el origen de nuevas especies. Se requiere algo más, que en la época de Darwin, o sea a mediados del siglo XIX, todavía se desconocía: por un lado, el mundo de la genética, y por otro, el pensamiento en términos de poblaciones, más allá que el individuo.
La evolución es el proceso de la transformación secuencial de las distintas formas de vida. La acumulación de cambios en una especie o una población es lo que se llama la microevolución, que se refiere a los procesos que hacen variar las proporciones de los alelos (variantes de los genes; ver mi post anterior) en una población. El mecanismo detrás de estos cambios es, a menudo, la selección natural o la selección sexual. La aparición de especies nuevas, o sea la especiación, el tema de este post, es lo que se conoce como la macroevolución: el fenómeno de que una especie dé origen a otra[ii]. Los dos términos se refieren a distintos niveles a que se puede observar la evolución; sin embargo, los procesos genéticos que dan origen a la macro y la microevolución son fundamentalmente los mismos[iii].
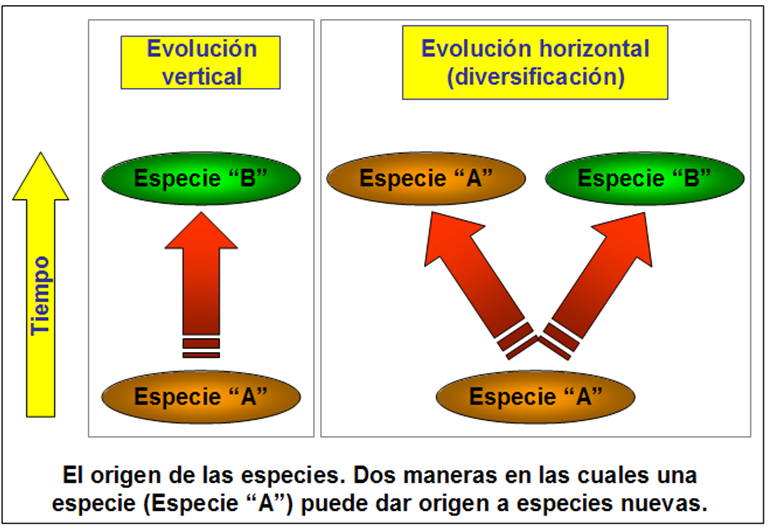
Existen dos maneras en la cual puede ocurrir la especiación: una se llama la evolución horizontal, y otra la evolución vertical. La siguiente figura ilustra estos dos tipos. En el caso de la evolución vertical, una especie se transforma hasta convertirse en otra especie – algo que en la naturaleza ocurre poco. Más frecuente es la evolución vertical, que es un proceso de diversificación en la cual una especie se separa en dos especies. En este proceso, de la especie original (llamémosla “Especie A”) puede ramificarse una especie nueva (“Especie B”), mientras que Especie A se mantiene sin cambios; es así que la corriente biológica de la cladística considera que se desarrolla la evolución[iv]. En este post estaremos considerando este tipo de evolución: la ramificación de una especie en dos.
¿Qué es una especie?
Para que podamos entendernos bien, es preciso definir el concepto de la especie. Existen unas cuantas definiciones de lo que es una especie[v]. Una de ellas, la morfológica, dice que si dos organismos son de la misma especie si tienen ciertos rasgos en común. Este concepto de la especie deja espacio para subjetividad: lo que una persona llamaría una sola especie con ciertas variantes, otra persona pudiera considerarlo dos especies muy parecidas. Además, se ha establecido que hay ocurrencias de especies que lucen casi idénticas, pero cuyo material genético es suficientemente diferente para considerarlas especies distintas. Sin embargo, en la paleontología, la ciencia que estudia especies de épocas pasadas de las cuales no nos queda nada sino restos fósiles (generalmente de huesos y/o dientes), estas definiciones morfológicas son las únicas que se pueden utilizar.
La definición biológica o zoológica, por otro lado, es más práctica para el estudio de especies actuales y por lo tanto la utilizaremos aquí. Esta definición se puede resumir de la siguiente manera: dos organismos pertenecen a la misma especie si pueden aparearse produciendo una progenie fértil. Esta definición es una variación de la definición original del biólogo Ernst Mayr, la cual reza que especies son “groups of interbreeding natural populations reproductively isolated from other such groups” [vi]. Esto implica que caballos y asnos son dos especies distintas, puesto que la mula, producto del apareamiento de un burro y una yegua, es infértil. Un perro doberman y un chihuahua, por otro lado, pertenecen a la misma especie puesto que – por lo menos en principio – pueden aparearse produciendo cachorros fértiles. La aplicabilidad de esta definición se limita a los animales, vegetales y hongos, dejando de lado a las bacterias[vii], pero aparte de esto, es ésta la definición más útil para nuestros fines.
Evolución por diversificación
La evolución por diversificación, o sea la ramificación de una especie desde otra, es un proceso de divergencia. La figura abajo muestra cómo ocurre este proceso. Básicamente se trata de un aumento de la diferencia genética entre dos poblaciones de una especie, que termina por volverse tan grande que los miembros de una población ya no son capaces de aparearse exitosamente con miembros de la otra: al presentarse esta situación, ya no se trata de dos poblaciones de una especie, sino de dos especies separadas[viii].
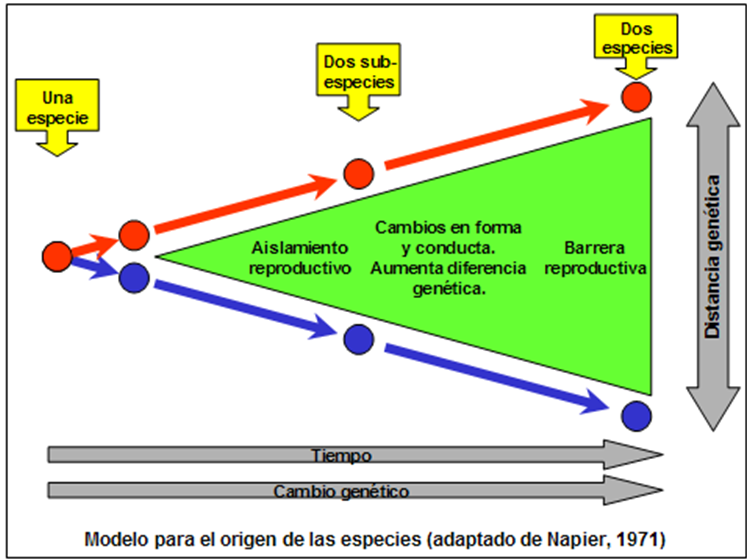
Regresemos al inicio de este proceso. Primero, veamos cómo se pueden formar dos poblaciones separadas de una especie. Para que éstas puedan seguir caminos evolutivos distintos, es preciso que exista algún tipo de barrera reproductiva entre las dos, para que no haya mezcla entre los dos grupos – puesto que, al haber mezcla, se desharía el proceso de diferenciación de los dos grupos. Abajo veremos qué tipos de barrera se han identificado.
Tal como ya indicó Darwin, aunque sin conocer el mecanismo, todas las especies tienden a cambiar en el tiempo. Ahora sabemos que esto es por cambios genéticos, que por pequeños que puedan parecer, al acumularse pueden resultar en cambios importantes en el organismo. Debido a la presencia de una barrera reproductiva entre las dos poblaciones, los cambios genéticos en una población van a ser distintos a los que experimenta la otra población, de manera que paulatinamente se dará una divergencia de las dos poblaciones. Cuando las diferencias en forma y conducta entre las dos son tales que, aun pudiendo aparearse, individuos de las dos poblaciones dejan de hacerlo, las dos pueden ser consideradas sub-especies de la especie a la cual ambas pertenecen.
La siguiente fase se da cuando, al aumentar la diferencia genética y la de forma y conducta entre las dos sub-especies, estas diferencias se vuelven tales que miembros de una ya no pueden aparearse con miembros de la otra produciendo una progenie fértil. En este momento ya no se habla de sub-especies, sino de especies propiamente dicho.
Aislamiento reproductivo
Tal como vimos arriba, para que se pueda dar especiación es necesario un aislamiento reproductivo de los grupos que van a dar origen a las especies nuevas. Si se da un aislamiento reproductivo entre dos poblaciones de una misma especie, ya no pueden parearse ejemplares de una población con ejemplares de la otra, de manera que un cambio genético que se da en una población no va a ser compartido con la otra. Como consecuencia, paulatinamente las dos poblaciones irán su propio camino evolutivo, adaptándose a sus respectivos entornos mediante selección natural[ix], y dado suficiente tiempo las dos poblaciones pueden convertirse en dos especies distintas. Pero es fundamental que se mantenga el aislamiento reproductivo durante suficiente tiempo, o que por lo menos se reduzca a su mínima expresión, para que no puedan volver a mezclarse las dos poblaciones mientras que sean todavía capaces de parearse y producir progenie fértil.
Obviamente, en la fase inicial de la divergencia entre dos poblaciones seguirá habiendo cruces entre las dos, resultando en un intercambio de material genético entre las dos poblaciones. Este flujo genético es inevitable. Por lo tanto, para que pueda haber especiación se requiere de unos mecanismos tan fuertes que sean capaces de contrarrestar el flujo genético que ocurre cuando dos poblaciones aun no estén aisladas reproductivamente. Se han identificado varios mecanismos, que pueden actuar solos o, mejor, conjuntamente, creando obstáculos para el flujo genético.
Aislamiento pre y post-cigótico
El proceso de especiación empieza por el desarrollo de un aislamiento reproductivo. Se hace una distinción entre aislamiento precigótico (una barrera que impide que se apareen ejemplares de distintas poblaciones) y postcigótico (cuando, si se cruzan ejemplares de poblaciones diferentes, los descendientes son estériles – tal como en el caso de la mula, mencionado arriba).
El aislamiento precigótico puede ser causado por separación en hábitat (si dos poblaciones viven en entornos distintos), separación temporal (si las poblaciones tienen horarios distintos, por ejemplo en el momento de aparearse), por barreras a la inmigración (si migrantes de una población a otra no logran reproducirse por no estar adaptados al nuevo entorno), por aislamiento sexual (si miembros de una población ya no están atraídos sexualmente por miembros de otra población; ver abajo), o por una barrera física que impida al semen de un macho llegar al aparato reproductor de una hembra de otra población (por incompatibilidad de órganos sexuales; volveremos a hablar de esto más abajo).
El aislamiento postcigótico puede deberse a incompatibilidad genética intrínseca (de manera que no hay descendencia, o que sí hay un descendiente híbrido pero éste se muere antes de reproducirse, o – tal como en el ejemplo de la mula – resulta estéril), a aislamiento ecológico (cuando el descendiente híbrido no está adaptado ni al entorno de la madre, ni al del padre), o a aislamiento sexual (cuando el híbrido no logra encontrar pareja, por ser distinto a ambas poblaciones de origen)[x].
Mutaciones
Mutaciones que aparecen en alguna población pueden reforzar las barreras reproductivas entre poblaciones, y a su vez se pueden beneficiar por estas barreras[xi]. Si en una población ocurre una coevolución sexual, en la que cambios en un sexo provocan cambios contrarios en el otro sexo, la velocidad de esta coevolución suele ser tal que la población alcanza rápidamente el aislamiento reproductivo[xii].
También se ha determinado que dos poblaciones de una misma especie pueden alcanzar un aislamiento reproductivo más fácilmente en una situación de estrés, por ejemplo debido a falta de alimento. En este tipo de condiciones la respuesta de la selección natural suele ser más fuerte: aparecen más mutaciones que pudieran ayudar a adaptar mejor las poblaciones a la escasez de alimentos, y entonces es más fácil que en una población empiece a predominar una mutación, y otra mutación en la otra población. Se observó que hasta con una sola mutación distinta entre las dos poblaciones puede surgir una barrera reproductiva[xiii].
Cabe destacar que el aislamiento reproductivo por sí solo no brinda seguridad de que vaya a haber especiación. Al parecer se requiere más que el solo aislamiento para que se desarrolle una especie nueva[xiv].
El aislamiento reproductivo, pues, es crítico, aunque no necesariamente suficiente, para que pueda darse especiación. Debe haber alguna barrera que impida que ocurran cruces entre ejemplares de los dos grupos. Veamos cuáles son los mecanismos que conocemos hasta la fecha para que se pueda erigir una barrera suficientemente efectiva.
Especiación alopátrida
El primer tipo de barrera reproductiva es de índole geográfica: si dos poblaciones son separadas por alguna barrera geográfica – por ejemplo un brazo de mar, una cadena montañosa, un río ancho –, es posible que ya no haya comunicación entre las mismas. Otro ejemplo es el de una población que se encuentra en una isla y otra en tierra firme, o en otra isla. En estos casos, las dos poblaciones se encuentran en ubicaciones claramente separadas, y la especiación que puede ocurrir bajo estas condiciones se llama especiación alopátrida (que tiene algunas variantes, llamadas peripátrida y parapátrida, que también se basan en una separación geográfica, aunque no tan completa, entre poblaciones). El gran biólogo Ernst Mayr (1904-2005) fue uno de los principales impulsores de la separación geográfica como la principal causa de especiación[xv].
Es más fácil separar geográficamente dos poblaciones de especies que no se muevan tanto, que poblaciones de especies con mayor movilidad. Se confirmó esto mediante un estudio de escarabajos: hay más especies de escarabajos no voladores que de escarabajos que sí vuelan[xvi]. Lo mismo se observó en el caso de aves tropicales: las que viven cerca del suelo, y que por lo tanto tienen menor movilidad que las especies que viven en los topes de los árboles, tienen una diversidad de especies mayor que las aves en los topes[xvii].
Sin embargo, una separación geográfica por si sola no conduce necesariamente a especiación. Por ejemplo, se determinó que poblaciones de anolis (un lagarto centroamericano) que habían estado separados por varios millones de años, viviendo en islas distintas del Caribe, no se habían convertido en especies distintas: eran capaces de cruzarse sin ningún inconveniente, produciendo descendientes perfectamente normales[xviii]. Al parecer, se requiere más para que se dé un desarrollo de especies nuevas.
Asimismo, la creación de grandes barreras tales como cadenas montañosas no siempre causa la creación de nuevas especies. El levantamiento de los Andes, por ejemplo, ayudó en el proceso de especiación, pero no fue el único factor en causar la aparición de especies nuevas[xix]. Veamos, pues, qué otros tipos de especiación existen.
Especiación sinpátrida
Pero no siempre la especiación se da cuando dos poblaciones se encuentran separadas geográficamente. Más bien se han observado más casos de especiación en poblaciones que se encuentran en una misma zona – lo que se llama especiación sinpátrida. ¿Cómo puede ocurrir eso?
Conocemos varios mecanismos por los cuales se puede dar especiación sinpátrida. He aquí algunos de los mejor conocidos, que pueden ocurrir bien sea separadamente, bien sea conjuntamente[xx].
Especiación ecológica
En primer lugar, dos poblaciones de una misma especie pueden empezar a ocupar nichos ecológicos distintos, o sea que aun viviendo en la misma área geográfica, ya casi no se encuentran más por haber desarrollado preferencias distintas acerca de ambientes o conductas. Esto se llama especiación ecológica, que por cierto puede ocurrir también en el caso de la especiación alopátrida[xxi].
El ejemplo clásico de esto son los pinzones de las islas Galápagos, ya mencionados al inicio de este post. En estas islas existen distintas especies de pinzones, cada una con un tamaño y una forma de pico diferente, adaptado a la fuente de alimentación del pinzón. Algunos pinzones tienen picos robustos, adaptados a abrir las cáscaras duras de semillas, otros los tienen más finos, adaptados para comer insectos, etcétera. Al adaptarse los pinzones a una forma de alimentarse específica, se generaron barreras reproductivas: se separaron en poblaciones distintas, cada una en su nicho ecológico, que aun compartiendo su isla con otros pinzones con otras adaptaciones alimenticias, ya no se cruzan con ellos. Este aislamiento reproductivo causó especiación ecológica[xxii]. Genéticamente, es un solo gen que controla la forma de los picos, de manera que la adaptación de los mismos a la alimentación de los pinzones fue fácil[xxiii].
Otro ejemplo es el caso de un pequeño pez llamado el espinoso, en un lago en Canadá. Allí ocurrió una especiación cuando, de dos poblaciones de una especie de espinoso, una empezó a vivir en aguas más profundas, alimentándose de zooplancton, mientras que otra se quedó en aguas someras, alimentándose de organismos bentónicos, o sea, que viven en el fondo lacustre. Al empezar a vivir en nichos ecológicos distintos, aunque en el mismo lago, las dos poblaciones de espinosos se separaron lo suficientemente como para que se originaran una barrera reproductiva y, después de múltiples generaciones, dos especies distintas.
Los corales (los organismos que forman arrecifes en las aguas cálidas) muestran un fenómeno parecido: se detectó cómo una especie se ha separado en dos especies distintas, cada una a una profundidad distinta[xxiv].
Dos variantes del caracol marino Littorina saxatilis, que vive en costas rocosas de España, parecen estar en vías de separarse en dos especies, ya que ocupan distintas posiciones en la zona costera y se están adaptando a sus entornos distintos, desarrollando en el camino una barrera reproductiva que por los momentos no está completa, pero que en un futuro lo pudiera ser[xxv].
Otro caso de una separación de dos poblaciones, al parecer sinpátrida, ha sido observado en ratas-topo ciegas en Israel: en una misma área ocurren dos formaciones rocosas distintas, cada una con su propio tipo de vegetación, y mientras que una población de ratas-topo ciegas vive sobre una de las dos formaciones rocosas, la segunda población vive sobre la otra formación. Datos genéticos indican que hay muy pocos cruces entre las dos poblaciones y que estas pudieran estar en vías de especiación[xxvi].

En el mundo vegetal, dos especies de mímulo (una mata americana con flores llamativos; ver figura) se separaron al desarrollar, mediante unos pocos cambios genéticos, primero colores distintos: una cambió el color de sus flores a rojo, que no atrae a las abejas mas sí a los colibríes, mientras que la otra se quedó de color rosa – un color que sí atrae a las abejas, pero espanta a los colibríes. Después desarrollaron formas distintas: la variante roja cambió la forma de su flor, alargándola de tal manera que sólo los colibríes, con sus picos largos, puedan alcanzar el néctar. De esta forma, las dos variantes de mímulo alcanzaron una barrera reproductiva entre las dos, ya que los animales que visitan a una no visitan a la otra, y vice versa; así las abejas polinizan sólo al mímulo rosa, y los colibríes sólo al rojo[xxvii].
Terminemos este recorrido por el mundo de la especiación ecológica considerando un caso de especiación reciente, observado en unas moscas de fruta (Rhagoletis). Las moscas de la fruta, que se encuentran en muchos laboratorios del mundo por ser útiles para estudios genéticos, provienen de Norteamérica. Son parásitos extremadamente especializados, que solamente se nutren de, y se aparean en, un único tipo de fruta. La especialización extrema de las moscas de la fruta crea barreras reproductivas fuertes. En este sentido, son un ejemplo perfecto de especiación ecológica.
Pero también hay especies de Rhagoletis en frutos que se importaron en Norteamérica hace pocos siglos: la manzana y la madreselva asiática, entre otros. La especie Rhagoletis pomonella es la que se especializó en las manzanas; se convirtió en uno de los primeros ejemplos conocidos de una especie que se originó por especiación sinpátrida[xxviii].
En los Estados Unidos se encontró en 1997 una nueva especie de Rhagoletis, en frutos de madreselva asiática. Puesto que la madreselva asiática fue importada en los Estados Unidos hace sólo unos 250 años, cabe la pregunta ¿de dónde se originó esa Rhagoletis de la madreselva?
La mosca fue sometida a estudios genéticos[xxix], y resultó que su material genético contiene algunos variantes que pertenecen a una especie de Rhagoletis que sólo vive en arándanos, pero también contiene otros variantes provenientes de una especie de Rhagoletis exclusiva de la baya de nieve. Se concluyó que la mosca de la madreselva se originó como producto del apareamiento entre una mosca de arándano y otra de baya de nieve (ambas especies son indígenas de América del Norte). Este proceso se llama hibridación; abajo hablaré más al respecto. La especialización extrema de las moscas de la fruta crea barreras reproductivas fuertes, lo que permite que – en el caso muy raro de que una mosca de la fruta haga el salto a otro tipo de fruta – puedan formarse especies nuevas mediante el proceso de especiación ecológica.
Pero el cuento de las moscas de la fruta no termina aquí. Al diversificarse, cuando empezan a infestar nuevas frutas y generan así nuevas especies (o variantes) especializadas, dan a su vez origen a ulteriores procesos de especiación. Resulta que las moscas de la fruta tienen a su vez un parásito, una pequeña avispa del género Diachasma. Cuando se forma una nueva especie de Rhagoletis, aislada de otras especies de sus congéneros, su avispa parasitaria también se queda aislada reproductivamente, y se origina una nueva especie de avispa. Este proceso se observó en el caso de una nueva especie de mosca de la fruta que se formó, especializada en las manzanas (la especie Rhagoletis pomonella, que ya vimos arriba), lo que causó el desarrollo de una nueva especie de avispa parasitaria especializada en esta nueva mosca, Rhagoletis pomonella[xxx].
Selección sexual
En segundo lugar, la selección sexual, de la cual hablamos en mi post anterior, puede jugar un papel importante en la especiación. La selección sexual ocurre cuando los machos de una especie empiezan a desarrollar distintos colores u otros rasgos característicos, y las hembras empiezan a desarrollar preferencias, unas por un patrón, otras por otro. De esta manera se forman múltiples grupos separados, cada uno conteniendo machos con unos colores o rasgos distintivos y hembras atraídas por estos. Al encontrarse en grupos separados que, los ejemplares de la especie no volverán a mezclarse, aun compartiendo el mismo entorno, de manera que pueden desarrollarse en especies distintas[xxxi]. Modelos indican que, especialmente en combinación con la selección natural (cuando dos poblaciones de una misma especie se adaptan a entornos distintos), la selección sexual es capaz de causar la aparición de nuevas especies[xxxii]. Sin que haya algún tipo de selección sexual, es muy difícil que haya especiación.
En cíclidos (peces parecidos a la perca) en el Lago Victoria, en África oriental, por ejemplo, se dio especiación causado por coloraciones distintas de los machos. Dentro de ciertas poblaciones, los machos empezaron a desarrollar colores distintos; esto puede ocurrir de manera fortuita, por deriva genética[xxxiii]. Cuando algunas hembras empezaron a preferir aparearse con estos machos de colores distintos, se originó una población distinta que, aun viviendo en el mismo entorno que los cíclidos originales, empezó a comportarse como un grupo aislado reproductivamente, por no aparearse con ejemplares de los grupos originales. Esto causó el desarrollo de no una, sino múltiples especies nuevas, caracterizadas por colores distintos a los de la especie original.
Volviendo a los espinosos, aquellos peces canadienses: se encontraron, también en Canadá, grupos de espinosos que desarrollaron barreras reproductivas por el hecho de que las hembras prefieren parejas que no sólo tengan los mismos colores, sino que también huelan como sus padres. Al generarse variantes de color y olor en la población, este proceso de impresión causó que poblaciones con colores y olores diferentes se convirtieron rápidamente en grupos aislados reproductivamente, ya que las hembras sólo se aparean con machos que son como sus papás[xxxiv].
El olor parece ser más importante de lo que se creía para distinguir poblaciones de una misma especie. Dos poblaciones de juncos (un pájaro norteamericano) en California emanan cada uno un conjunto de compuestos químicos distintos, lo que pudiera contribuir a una mayor separación entre las dos poblaciones y, en algún momento, una separación en dos especies diferentes[xxxv].
En ciertos casos, la selección sexual lo es de manera muy literal: se trata de especies, por ejemplo de insectos, que se parecen mucho y viven en las mismas áreas, pero que están separadas por una barrera reproductiva formidable: tienen órganos sexuales de formas diferentes, de manera que solamente pueden aparearse con miembros de su propia especie. Este modelo para la aparición de aislamiento reproductivo, llamado “llave y cerradura”, fue propuesto ya en el siglo XIX. Sin embargo, es sólo hace unos años que se pudo determinar cuál es el mecanismo que puede causar cambios en el tamaño y/o la forma de los órganos copulatorios – órganos que por su función no suelen variar mucho de forma o tamaño. El mecanismo hallado, en el caso de unas especies de escarabajos, es que existe una relación inversa entre el tamaño del órgano sexual de los machos y el tamaño de sus cuernos: si los cuernos son grandes, al parecer no queda mucha posibilidad para que crezca mucho el órgano sexual, mientras que si los cuernos son pequeños, el órgano sexual puede crecer más y alcanzar unos tamaños relativamente enormes. Obviamente, los machos con órganos sexuales grandes suelen aparearse con otras hembras que los machos cuyos órganos sexuales son modestos, lo que crea barreras reproductivas entre poblaciones y especies. Resulta que el tamaño de los cuernos es una adaptación al entorno de los escarabajos: si la densidad poblacional de los escarabajos es baja, los machos suelen ser más agresivos y requieren de cuernos grandes, lo que va a costa del tamaño de sus órganos sexuales; mientras que en el caso de una densidad poblacional alta, los machos son menos agresivos, lo que se refleja en cuernos más modestos y por ende, órganos sexuales grandes[xxxvi].
Al parecer, la selección sexual es un factor importante en el proceso de especiación. Pero, según ciertos modelos, por sí solo no resulta fácilmente en especiación: también se requiere un factor tal como una bipolaridad de fuentes de alimento, o sea, recursos bien distintos que permiten a ejemplares especializarse en una fuente de alimentación, o en otra – lo que, tal como vimos arriba, es un caso que puede llevar a selección ecológica. En otras palabras, especiación sinpátrida puede ocurrir cuando son dadas ciertas condiciones conjuntamente: por ejemplo, selección sexual combinada con selección ecológica[xxxvii].
Hibridación
En tercer lugar, en el reino vegetal y, más raramente, en el animal, aparecen de vez en cuando nuevas especies por los procesos llamados hibridación y poliploidía. La hibridación ocurre cuando un ejemplar nace como resultado de un cruce entre dos ejemplares de especies distintas. En el mundo animal, un ejemplo de hibridación es la mula, que tal como ya mencionamos es el fruto del cruce entre una yegua y un burro. Las mulas no son fértiles, pero en el mundo vegetal, un híbrido sí puede conservar cierta fertilidad, aunque reducida. A veces el híbrido es lo suficientemente distinto de cada uno de sus progenitores que puede dar origen a una especie distinta (muchas plantas no necesitan de otro ejemplar de la misma especie para reproducirse).
En el mundo animal la creación de nuevas especies por hibridación es muy rara, mas no imposible. Prueba de ello son especies de mariposas de las cuales se pudo comprobar que se originaron por hibridación, o sea por un cruce entre dos ejemplares de especies distintas. Puesto que las mariposas híbridas tienen en sus alas patrones de colores distintos a los de sus progenitores, difícilmente van a encontrar parejas con quienes aparearse. Pero si logran hacerlo, se puede formar una especie nueva, aislada genéticamente de ambas especies ancestrales por barreras reproductivas debido a sus colores distintos[xxxviii].
La poliploidía es un fenómeno genético. De cada organismo conocido el material genético consiste de cierta cantidad de cromosomas: estructuras en las células que contienen el ADN del organismo. Normalmente, los individuos que nacen por reproducción sexual tienen dos juegos completos de cromosomas: uno de cada uno de los progenitores. En el caso de poliploidía, el individuo recibe de sus progenitores no dos, sino tres o más juegos completos de cromosomas; y si los progenitores pertenecen a dos especies distintas, el individuo resultante sería tanto poliploide como híbrido[xxxix]. La poliploidía es bastante común entre las plantas. Un ejemplar poliploide difícilmente puede aparearse exitosamente con otros ejemplares que no sean poliploides (o no lo sean de la misma manera), y así, tal como vimos en el caso de hibridación, puede dar origen a una nueva especie, por lo menos logra reproducirse.
Un ejemplo de poliploidía es el de otra especie de mímulo. Esta especie fue introducida en las islas británicas alrededor del 1800. Recientemente se descubrió en Escocia una especie de mímulo indígena, a la cual fue dado el nombre de Mimulus peregrinus. Un estudio genético evidenció que se originó como híbrido poliploide a partir de dos otras especies de mímulo, ambas importadas en el siglo XIX. A diferencia de otros híbridos, esta especie es altamente fértil, y el hecho que no se ha expandido mucho sugiere que se formó hace poco tiempo[xl].
Especiación cromosómica
Por último, se ha propuesto como posible mecanismo de especiación, la especiación cromosómica, o sea especiación por reordenamiento cromosómico. Un reordenamiento cromosómico ocurre si una parte de una cromosoma se invierte o mueve. Cuando esto se dé, la parte reordenada de la cromosoma no participa en el proceso de recombinación del material genético cuando se fusionan un óvulo y un espermatozoide: se mantiene como una “isla”, que pasa inalterada y entera a la progenie. Normalmente, esto causaría una progenie inviable o infértil; en ciertos casos, sin embargo, el ejemplar logra sobrevivir aun con un reordenamiento cromosómico. De esta manera, el reordenamiento cromosómico puede ayudar a crear una barrera reproductiva. Además, el material genético reordenado, al estar aislado, puede acumular mutaciones más rápidamente que el resto del genoma, de manera que la población en la cual ocurre puede cambiar rápidamente hasta convertirse en una especie distinta[xli].
La especiación cromosómica es un mecanismo capaz de contrarrestar el flujo genético que ocurre cuando dos poblaciones aun no están aisladas reproductivamente, y todavía ocurren cruces entre las dos.
Conclusión
El desarrollo de nuevas especies, la especiación, empieza por la aparición de barreras reproductivas entre las poblaciones de una especie, para que estas puedan diversificarse y convertirse en especies distintas. Se han identificado múltiples mecanismos que puedan formar tales barreras, dando así origen a un aislamiento reproductivo.
Tradicionalmente, se consideró que la principal barrera reproductiva es la geográfica, y que la especiación es principalmente alopátrida. Sin embargo, más recientemente se ha determinado que la especiación a menudo es sinpátrida, o sea que puede ocurrir aun cuando dos poblaciones estén viviendo en la misma área geográfica. Los principales mecanismos para un aislamiento reproductivo entre especies que comparten el mismo espacio son:
- Especiación ecológica. Cuando dos poblaciones de una misma especie ocupan nichos ecológicos distintos, puede crearse una barrera reproductiva entre las dos.
- Selección sexual. Cuando en una población empiezan a aparecer machos con colores u otros rasgos distintos, y ciertas hembras prefieren aparearse con estos en lugar de los machos “tradicionales”, puede formarse un nuevo grupo, aislado reproductivamente.
- Hibridación. Cuando un ejemplar nace como resultado de un cruce entre dos ejemplares de especies distintas, puede ser lo suficientemente distinto de ambas especies ancestrales para no ser capaz de aparearse con ninguna de las dos, dando potencialmente origen a un linaje nuevo.
- Especiación cromosómica. Cuando ocurre un reordenamiento cromosómico y el ejemplar con tal condición ya no logra obtener descendencia con sus congéneros.
La especiación ecológica y la selección sexual resultan en un aislamiento precigótico, o sea, impiden que haya apareamiento entre dos ejemplares de distintas poblaciones o especies. También es posible que se dé un aislamiento precigótico por razones físicas, cuando el órgano sexual masculino no logra acoplarse con el femenino. En el caso de la hibridación y la especiación cromosómica, el aislamiento es más bien postcigótico, o sea, puede ocurrir un apareamiento, pero la descendencia no es viable, o sale estéril.
De estos mecanismos, ninguno suele actuar por sí solo en el proceso de especiación. Generalmente, actúan en conjunto dos o más mecanismos: por ejemplo, la selección sexual actúa conjuntamente con la especiación ecológica, o con la hibridación.
De esta manera se pudo solucionar el enigma que no logró resolver Darwin, cuando se dio cuenta de que su teoría de la selección natural no era suficiente para explicar la aparición de nuevas especies. Ya sabemos cómo se originaron las distintas especies de pinzones que Darwin encontró en las islas Galápagos.
Nota: partes de este post fueron publicadas originalmente en mi blog – ahora cerrado – “Tiempos de Cambio”. Los dibujos en el post fueron elaboradas por el autor. La imagen que acompaña al título es de pinzones de Darwin, de las islas Galápagos, tomada de la descripción que Darwin publicó de su viaje en el Beagle (fuente: http://commons.wikimedia.org/wiki/File:Darwin%27s_finches_by_Gould.jpg).
[i]. Darwin, Ch., 1859/1968. The origin of species by means of natural selection. Penguin Books, Londres.
[ii]. Zimmer, C., 2008. What is a species? Scientific American, junio 2008, 72-79. www.sciam.com. Ver también: http://es.wikipedia.org/wiki/Macroevoluci%C3%B3n.
[iii]. Wilkins, J., 2006. Macroevolution. Its definition, philosophy and history. Disponible en: http://www.talkorigins.org/faqs/macroevolution.html.
[iv]. El concepto del cladismo (mas no su nombre) fue desarrollado a mediados del siglo XX por el biólogo alemán Willi Hennig: Hennig, W., 1966. Phylogenetic systematics. University of Illinois Press, Chicago.
[v]. Ver: http://es.wikipedia.org/wiki/Especie.
[vi]. Mayr, E., 1942. Systematics and the origin of species from the viewpoint of a zoologist. Colombia University Press, New York.
[vii]. Margulis, L. y Sagan, D., 2002. Acquiring genomes. A theory of the origin of species. Basic Books, New York.
[viii]. Ver: http://es.wikipedia.org/wiki/Especiaci%C3%B3n.
[ix]. Funk, D.J., Nosil, P. y Etges, W.J., 2006. Ecological divergence exhibits consistently positive associations with reproductive isolation across disparate taxa. Proceedings National Academy of Sciences, 103 (9), 3209-3213. www.pnas.org.
[x]. Rundle, H.D. y Nosil, P., 2005. Ecological speciation. Ecology letters, 8, 336-352. http://onlinelibrary.wiley.com/doi/10.1111/j.1461-0248.2004.00715.x/pdf.
[xi]. Barton, N.H. y Rodríguez de Cara, M.A., 2009. The evolution of strong reproductive isolation. Evolution, 63 (5), 1171-1190. http://onlinelibrary.wiley.com/doi/10.1111/j.1558-5646.2009.00622.x/pdf.
[xii]. Martin, O.Y. y Hosken, D.J., 2003. The evolution of reproductive isolation through sexual conflict. Nature, 423, 979-982. http://www.nature.com/nature/journal/v423/n6943/full/nature01752.HTML.
[xiii]. Kroll, E., Coyle, S., Dunn, B., Koniges, G., Aragon, A., Edwards, J. y Rosenzweig, F., 2013. Starvation-associated genome restructuring can lead to reproductive isolation in yeast. PLoS ONE, 8 (7), e66414. www.plosone.org.
[xiv]. Rabosky, D.L. y Matute, D.R., 2013. Macroevolutionary speciation rates are decoupled from the evolution of intrinsic reproductive isolation in Drosophila and birds. Proceedings National Academy of Sciences, 110 (38), 15354-13359. http://www.pnas.org/content/110/38/15354.full.pdf+html?with-ds=yes.
[xv]. Mayr, 1942. Ver nota 6. Ver también: http://en.wikipedia.org/wiki/Allopatric_speciation; http://www.the-scientist.com/?articles.view/articleNo/15228/title/Mechanisms-of-Speciation/.
[xvi]. Ikeda, H., Nishikawa, M. y Sota, T., 2012. Loss of flight promotes beetle diversification. Nature Communications, 3, 648. http://www.nature.com/ncomms/journal/v3/n1/pdf/ncomms1659.pdf.
[xvii]. Tilston Smith, B., y otros, 2014. The drivers of tropical speciation. Nature, 515, 406-409. http://www.nature.com/nature/journal/v515/n7527/full/nature13687.html.
[xviii]. Thorpe, R.S., Surget-Groba, Y. y Johansson, H., 2010. Genetic tests for ecological and allopatric speciation in anoles on an island archipelago. PLoS Genetics, 6 (4), e1000929. www.plosgenetics.org.
[xix]. Tilston Smith y otros, 2014. Ver nota anterior.
[xx]. Albert, A.Y.K. y Schluter, D., 2005. Selection and the origin of species. Current Biology, 15 (8), R283-R288. http://www.sciencedirect.com/science/article/pii/S0960982205003866.
[xxi]. Rundle y Nosil, 2005. Ver nota 12.
[xxii]. Grant, P.R. y Grant, B.R., 2011. How and Why Species Multiply: The Radiation of Darwin’s Finches. Princeton Series in Evolutionary Biolgoy. Princeton University Press.
[xxiii]. Lamichhaney, S. y otros, 2015. Evolution of Darwin’s finches and their beaks revealed by genome sequencing. Nature, 518, 371-375. http://www.nature.com/nature/journal/v518/n7539/full/nature14181.html.
[xxiv]. Prada, C. yHellberg, M.E., 2013. Long prereproductive selection and divergence by depth in a Caribbean candelabrum coral. Proceedings National Academy of Sciences, 110 (10), 3961-3966. http://www.pnas.org/content/110/10/3961.full.pdf+html. Pfennig, D..W., 2013. Inviable immigrants drive diversification in the sea. Proceedings National Academy of Sciences, 110 (10), 3713-3714. http://www.pnas.org/content/110/10/3713.full.pdf+html.
[xxv]. Rolán-Alvarez, E., 2007. Sympatric speciation as a by-product of ecological adaptation in the Galician Littorina saxatilis hybrid zone. Journal of Molluscan Studies, 73, 1-10. http://mollus.oxfordjournals.org/content/73/1/1.full.pdf+HTML.
[xxvi]. Hadid, Y. y otros, 2013. Possible incipient sympatric ecological speciation in blind mole rats (Spalax). Proceedings National Academy of Sciences, 110 (7), 2587-2592. http://www.pnas.org/content/110/7/2587.full.pdf+html.
[xxvii]. Schemske, D.W. y Bradshaw, H.D., 1999. Pollinator preference and the evolution of floral traits in monkeyflowers (Mimulus). Proceedings National Academy of Sciences, 96 (21), 11910-11915. http://www.pnas.org/content/96/21/11910.full.pdf+html. Charlesworth, B. y Charlesworth, D., 2000. Reproductive isolation: Natural selection at work. Current Biology, 10 (2), R68-R70. http://www.sciencedirect.com/science/article/pii/S0960982200002803.
[xxviii]. Bush, G.L., 1969. Sympatric host race formation and speciation in frugivorous flies of the genus Rhagoletis (Diptera, Tephritidae). Evolution, 23 (2), 237-251. http://www.jstor.org/discover/10.2307/2406788.
[xxix]. Schwarz, D., Matta, B.M., Shakir-Botteri, N.L. y McPheron, B.A., 2005. Host shift to an invasive plant triggers rapid animal hybrid speciation. Nature, 436, 546-549. http://www.nature.com/nature/journal/v436/n7050/full/nature03800.HTML.
[xxx]. Forbes, A.A., Powell, T.H.Q., Stelinski, L.L., Smith, J.J. y Feder, J.L., 2009. Sequential sympatric speciation across trophic levels. Science, 323, 776-779. http://www.sciencemag.org/content/323/5915/776.full.pdf.
[xxxi]. M’Gonigle, L.K., Mazzucco, R., Otto, S.P. y Dieckmann, U., 2012. Sexual selection enables long-term coexistence despite ecological equivalence. Nature, 484 (7395), 506-509. http://www.nature.com/nature/journal/v484/n7395/full/nature10971.html.
[xxxii]. Van Doorn, G.S., Edelaar, P. y Weissing, F.J., 2009. On the origin of species by natural and sexual selection. Science, 326 (5960), 1704-1707. http://www.sciencemag.org/content/326/5960/1704.short.
[xxxiii]. Dieckmann, U. y Doebeli, M., 1999. On the origin of species by sympatric speciation. Nature, 400, 354-357. http://www.nature.com/nature/journal/v400/n6742/abs/400354a0.html.
[xxxiv]. Kozak, G.M., Head, M.L. y Boughman, J.W., 2011. Sexual imprinting on ecologically divergent traits leads to sexual isolation in sticklebacks. Proceedings Royal Society B, 278, 2604-2610. http://rspb.royalsocietypublishing.org/content/royprsb/278/1718/2604.full.pdf.
[xxxv]. Whittaker, D.J., Soini, H.A., Atwell, J.W., Hollars, C., Novotny, M.V. y Ketterson, E.D., 2010. Songbird chemosignals: volatile compounds in preen gland secretions vary among individuals, sexes, and populations. Behavioral Ecology, 21 (3), 608-614. http://beheco.oxfordjournals.org/content/21/3/608.full.pdf+html.
[xxxvi]. Parzer, H.F. y Moczek, A.P., 2008. Rapid antagonistic coevolution between primary and secondary sexual characters in horned beetles. Evolution, 62 (9), 2423-2428. http://onlinelibrary.wiley.com/doi/10.1111/j.1558-5646.2008.00448.x/abstract.
[xxxvii]. Thibert, Plante, X. y Hendry, A.P., 2011. Factors influencing progress toward sympatric speciation. Journal of Evolutionary Biology, 24, 2186-2196. http://onlinelibrary.wiley.com/doi/10.1111/j.1420-9101.2011.02348.x/full.
[xxxviii]. Mavárez, J., Salazar, C.A., Bermingham, E., Salcedo, C., Jiggins, C.D. y Linares, M., 2006. Speciation by hybridization in Heliconius butterflies. Nature, 441, 868-871. http://www.nature.com/nature/journal/v441/n7095/full/nature04738.html.
[xxxix]. Ver: http://es.wikipedia.org/wiki/Poliploid%C3%ADa.
[xl]. Vallejo-Marín, M., 2012. Mimulus peregrinus (Phrymaceae): A new British allopolyploid species. PhytoKeys, 14, 1-14. http://phytokeys.pensoft.net/articles.php?id=1432.
[xli]. Rieseberg, L.H., 2001. Chromosomal rearrangements and speciation. TRENDS in Ecology and Evolution, 16 (7), 351-358. http://www.sciencedirect.com/science/article/pii/S0169534701021875. Faria, R. y Navarro, A., 2010. Chromosomal speciation revisited: rearranging theory with pieces of evidence. TRENDS in Ecology and Evolution, 25 (11), 660-669. http://www.sciencedirect.com/science/article/pii/S0169534710001795. Faria, R., Neto, S., Noor, M. y Navarro, A., 2011. Role of natural selection in chromosomal speciation. En: Encyclopedia of Life Sciences (ELS). John Wiley & Sons, Ltd. http://onlinelibrary.wiley.com/doi/10.1002/9780470015902.a0022850/abstract.

{kind=link}
Saludos Peter, quisiera preguntarte sobre otro tema de interés, no sé qué tanto se ha verificado esta información, te dejo el link: https://noticiasdelaciencia.com/art/9064/recibir-por-herencia-genetica-un-conocimiento-que-adquirio-el-padre-o-la-madre
Gracias nuevamente por tus comentarios!
Me gustaMe gusta
Puede tratarse de transmisión epigenética. Se está viendo que cambios epigenéticos, que se dan durante la vida de un organismo (y que afectan cómo el ADN se expresa), pueden ser transmitidos ocasionalmente a la siguiente generación.
Me gustaMe gusta