
En el post del 26 octubre 2014 sobre el naturalista inglés Charles Darwin vimos cómo éste propuso, en 1859[1], el proceso de selección natural para explicar cómo pueden cambiar en el tiempo los seres vivientes. Este proceso es el que, hasta el día de hoy, forma la base de la teoría de la evolución.
En el presente post consideraremos en un poco más detalle cómo funciona la selección natural, y nos preguntaremos cuáles otras formas de selección existen.
Cómo funciona la selección natural
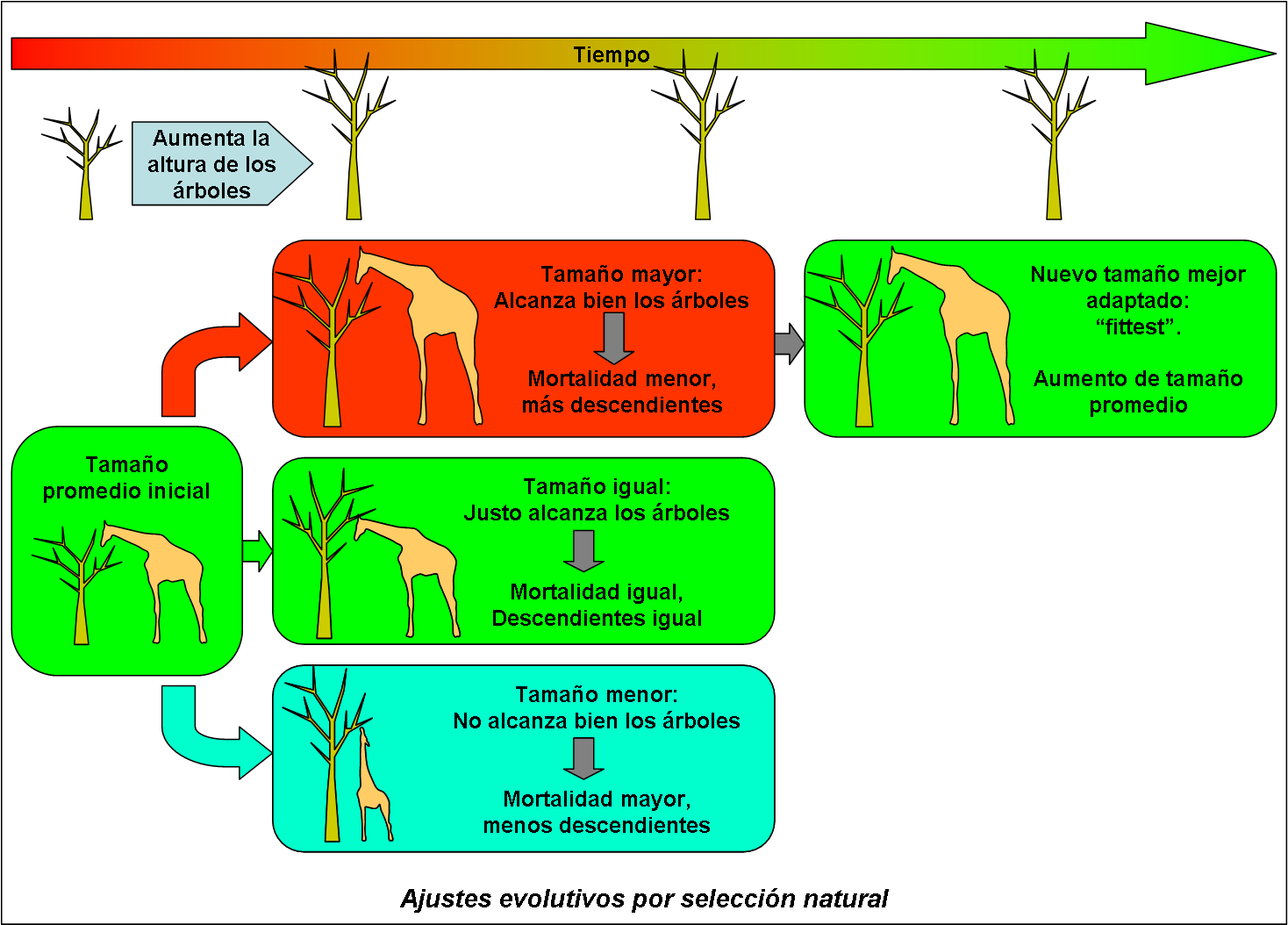
Para ver cómo funciona la selección natural, tomaremos como ejemplo las jirafas, animales que gracias a su cuello largo son capaces de comer las hojas de árboles altos que otros animales herbívoros no pueden alcanzar. Veamos la figura que acompaña este texto. Allí se muestra esquemáticamente cómo cambia el tamaño de las jirafas en el tiempo, a raíz de un cambio en la altura de los árboles de los cuales se nutren.
Supongamos que, por una razón u otra, empieza a aumentar la altura de estos árboles. Esto hará que aquellas jirafas cuyo tamaño está cerca del promedio de la especie, tendrán mayor dificultad en alcanzar el tope de los árboles, ahora más altos, pero aun lograrán alcanzar las ramas bajas y sobrevivir. Aquellos ejemplares cuya estatura está por debajo de lo normal, sin embargo, no lograrán alcanzar bien ni las ramas más bajas y les costará sobrevivir. Por otro lado, los ejemplares más altos de lo normal se encontrarán en una posición favorecida, puesto que serán los únicos en alcanzar hasta los topes de los árboles, obteniendo así suficiente comida aun en épocas de escasez.
Obviamente, las jirafas más altas de lo normal son las que “ganarán” esta carrera. Podrán vivir más tiempo y producir más descendientes, quienes – siguiendo las leyes de la genética – tenderán a ser relativamente altos también y a su vez producir más descendientes que aquellas jirafas que se quedaron cortas. Así aumentará en la población de jirafas el porcentaje de ejemplares altos, en detrimento de los ejemplares bajos, y el tamaño promedio de la población aumentará.
Este aumento de la estatura de las jirafas es un ejemplo de un cambio evolutivo por selección natural (natural selection). La palabra “selección” se refiere al hecho de que las nuevas condiciones del entorno de las jirafas favorecieron a los ejemplares mejor adaptados. La palabra “natural” indica que es el mismo entorno natural el que causa el cambio (abajo veremos algunas otras formas de selección).
En nuestro ejemplo, las jirafas altas resultaron ser las mejor adaptadas a la nueva situación, cuando aumentó la altura de los árboles, y por lo tanto las que mejor sobrevivieron. Esto se conoce como survival of the fittest (supervivencia del mejor adaptado). Este eslogan fue lanzado, no por Darwin, sino por un contemporáneo suyo, el filósofo inglés Herbert Spencer (1820-1903), quien también popularizó el término evolución[2]. Se define el fittest, el mejor adaptado (mejor adecuado), como aquel individuo en una población que más descendientes tenga. La adecuación (fitness) de un individuo es una medida de la capacidad del individuo de reproducirse.
Selección y genes
Cada organismo lleva, en forma de su material genético, su “plano de construcción”, con todas las instrucciones necesarias para su formación y desarrollo. Estas instrucciones están codificadas en los genes, de los cuales cada especie tiene hasta decenas de miles. Cada gen proporciona el código para alguna actividad, por ejemplo la elaboración de encimas que, a su vez, construyen y mantienen las células del organismo. En una población de cualquier especie animal o vegetal, los individuos tienen prácticamente los mismos genes. Pero siempre hay variaciones entre los genes de distintos individuos, ya que muchos genes vienen en variantes, llamados alelos. Por ejemplo, el gen que codifica para el color de nuestros ojos tiene varios alelos: uno que codifica para ojos marrones, otro para ojos azules, etcétera. Dependiendo de cuál alelo llevamos, nuestros ojos serán de un color o de otro. (Estoy simplificando el asunto; entre otras cosas, en la realidad no es un solo gen el que controla el color de los ojos.)
En el caso de muchos genes, no importa tanto cuál alelo está presente en el organismo. El color de los ojos, por ejemplo, no influye en la adecuación del organismo: un individuo de ojos azules puede tener tantos descendientes como uno de ojos verdes (a no ser que, culturalmente, una sociedad prefiera cierto color de ojos[3] – esto sería un ejemplo de selección cultural, de la que hablaremos en otra oportunidad). Pero en el caso de muchos otros genes, sí importa qué alelos el organismo tiene. Si el alelo fortalece al individuo, de manera que se vuelva más longevo y/o más fértil, y por lo tanto pueda tener más descendientes (o sea, ser más adecuado), este alelo va a estar sujeto a selección positiva: al causar un aumento de los descendientes, todos portadores del alelo, este alelo se convertirá – después de unas generaciones – en el más común en la población. En ese momento se puede decir que ocurrió evolución de la especie, por selección natural. Es así como cambios genéticos subyacen a los cambios evolutivos de una especie.
Si, por otro lado, el alelo vuelve más débil al organismo, ocurre lo opuesto: el individuo tendrá menos descendientes, de manera que el alelo en cuestión no se extenderá en la población, hasta prácticamente desaparecer. Esto se llama selección natural negativa: la eliminación de rasgos perjudiciales.
Resumiendo: el proceso de la selección natural es impulsado por mutaciones genéticas. Estas pueden resultar en un cambio en la forma o conducta del organismo (en lenguaje científico, la forma de un organismo se llama el fenotipo, mientras que su material genético se llama el genotipo). Los cambios pueden ser positivos (cuando aparece un alelo beneficioso), en el sentido de que aumenta la probabilidad de que el organismo logre procrearse, o sea de pasar su material genético a una siguiente generación; en el ejemplo de las jirafas que acabamos de ver, los cambios que resultan en jirafas más altas, obviamente son positivos. Cambios negativos ocurren también (cuando aparece un alelo perjudicial), pero la probabilidad de que un organismo con una mutación perjudicial logre procrearse es pequeña, así que tales cambios suelen desaparecer rápidamente de una población.
Muchos cambios son neutros: no resultan directamente en cambios en el fenotipo. Sí pueden contribuir a cambios futuros: por ejemplo, si un cierto cambio en el fenotipo requiere de una mutación del gen A y otra del gen B para que se pueda dar, una mutación de solamente el gen A no va a causar este cambio hasta que no se dé la mutación del gen B también.
En la realidad, la situación de los alelos es más compleja que la arriba descrita. Ya que, en el caso de reproducción sexual, todo individuo tiene dos progenitores, de cada uno de los que hereda un juego de genes, este tiene dos copias de prácticamente todos los genes. Las dos copias pueden ser idénticas o, en el caso de genes con variantes (o sea, alelos), diferentes. Por ejemplo, el individuo puede haber recibido del padre un alelo para ojos marrones, y de la madre un alelo para ojos verdes. Uno de los dos dominará, y determinará de cuál de los padres el individuo tomará el color de los ojos.
En el caso de un gen del que existen alelos beneficiosos y otros que son neutros o, inclusive, perjudiciales, es posible que un individuo recibe de uno de los padres un alelo beneficioso, y de otro, un alelo perjudicial. Generalmente no pasa nada: el individuo vive normalmente, gracias al alelo beneficioso. Pero en el caso de que el individuo reciba de ambos padres un alelo perjudicial, el organismo se va a ver afectado. En casos extremos, un individuo con dos alelos perjudiciales de un gen importante puede llegar a fallecer antes de procrearse – y así se reduciría la ocurrencia del alelo defectuoso en la población.
Una variante de selección natural en genes se llama “selección equilibradora” (balancing selection). El caso de la selección equilibradora se da cuando un alelo A de algún gen es beneficioso cuando el portador de ese gen sólo tiene una copia del alelo A; pero cuando ambos ejemplares del gen son del alelo A, el organismo puede verse afectado seriamente[4]. Un ejemplo clásico de esta situación es la anemia de células falciformes, una enfermedad que se debe a un mal funcionamiento de los glóbulos rojos en la sangre y generalmente resulta en la muerte precoz de la persona enferma. La enfermedad, que es genética, se produce cuando el individuo recibe de ambos padres un particular alelo de un gen que controla la forma de los glóbulos rojos. Se pudiera suponer que, de ser así, este alelo perjudicial desaparecería rápidamente de la población. Pero no es así, ya que el alelo, que es perjudicial para los que tienen dos copias del mismo, es beneficioso para aquellas personas que llevan una sola copia del mismo: estas personas tienen mayor resistencia a la malaria. Así que, por lo menos en aquellas regiones donde ocurre la malaria, es bastante común el alelo que brinda resistencia a esta enfermedad, aunque a un precio alto: la de una muerte precoz para aquellas personas que recibieron en el momento de su concepción no una, sino dos copias del alelo anti-malaria[5].
Otros tipos de selección. Selección sexual
Si Darwin lanzó el término “selección natural”, ¿habría otros tipos de selección también? Seguro, los hay.
Darwin utilizó como analogía del proceso de selección natural, el de “selección bajo domesticación” (selection under domestication). Este último proceso es impulsado por el hombre: por ejemplo el criador de perros, quien selecciona los perros que tienen alguna característica deseada y utiliza sólo aquellos para obtener crías, de manera que sus perros cambien de una manera específica, según las preferencias del criador.
Otro tipo de selección es la “selección sexual” (sexual selection). Este término proviene de Darwin también[6]. Lo acuñó para explicar la existencia de rasgos extravagantes y aparentemente gravosos, tales como la cola enorme y llamativa del pavo real o los grandes cuernos de ciertos venados machos. Concluyó que este tipo de rasgos sirven a los machos para atraer a hembras con quienes parearse y así asegurarse de una descendencia cuantiosa. Posteriormente se afinó esta explicación, planteando que estos rasgos extravagantes son una señal de fuerza y salud del macho, por lo que las hembras pueden determinar con cuáles de ellos parearse. Esto es lo que el biólogo israelí Amotz Zahavi llamó el “principio del handicap” (the handicap principle)[7].
El principio del handicap plantea que los rasgos extravagantes, costosos en términos de requerimientos energéticos o de exposición a riesgos, son una señal honesta que refleja la salud del macho, ya que individuos débiles no son capaces de lucirlos. Probablemente la señal no es completamente honesta, y habrá pavos reales débiles con colas impresionantes[8], pero por lo general los rasgos extravagantes corresponden con machos sanos y fuertes. Esto ha sido demostrado, por ejemplo, en el caso de los cuernos de ciervos machos[9].
El mecanismo de la selección sexual ha sido utilizado para explicar muchas de las diferencias que existen, en todo el reino animal, entre los dos sexos[10]. Esto aplica no sólo a los animales actuales, sino también a los prehistóricos, muchos de los cuales muestran diferencias entre los sexos (por ejemplo, crestas en la cabeza o el cuerpo) que pueden ser atribuidas a selección sexual; es inclusive posible que las plumas se originaran por esta razón, no para volar[11]. Asimismo, es probable que diferencias entre el tamaño de los picos de ciertas especies de gorriones no se originaron por diferencias en dieta, sino por selección sexual: las hembras prefieren parejas con picos grandes, tal vez por ser machos más viejos o sea más expertos y probados[12].
Entre las diferencias entre los sexos también hay diferencias en comportamiento. Una afirmación “clásica” en este sentido es la que plantea que los machos buscan el mayor número posible de parejas para así maximizar su descendencia[13], mientras que las hembras prefieren un número limitado de parejas, que más bien ayuden a cuidar los descendientes. Esta afirmación, sin embargo, se fundamenta en un estudio clásico utilizando la mosca Drosophila, que resulta tener una falla lógica, por lo que su conclusión pudiera no ser válida[14].
Al nivel de los genes, la selección natural no es el único proceso que determina cuáles mutaciones persistirán en el tiempo, y cuáles no. Procesos moleculares tales como la conversión sesgada de genes (biased gene conversion) pueden fijar las mutaciones genéticas en un organismo, de manera que tienen mayores probabilidades de ser transmitidas a sus descendientes, independientemente de si son mutaciones positivas o negativas, o sea al margen del proceso de selección natural[15].
En el caso de la selección sexual, el desarrollo de los rasgos extravagantes se debe a mutaciones genéticas que tienen expresión sólo en el caso de los machos. Esto hace posible que estos rasgos cambien rápidamente, o inclusive desaparezcan, dependiendo de si las ventajas que confieren los rasgos compensan, o no, los costes que los rasgos conllevan[16].
Niveles de selección. Selección de grupos, selección familiar
Hay controversia acerca de los niveles a los cuales actúa la selección natural. La ortodoxia biológica sostiene que la selección natural actúa sobre el individuo. Las mutaciones genéticas que permiten cambios en el organismo del individuo, los que la selección natural puede conservar si son positivos o neutros, y desechar si son negativos, se dan por definición en un solo individuo, que después puede pasar sus genes, mutaciones incluidas, a sus descendientes.
Algunos consideran que si el fenotipo (el organismo) no es sino un reflejo de lo que ocurre en el genotipo (genoma) del organismo, el organismo se puede entonces considerar como meramente el contenedor en el que se encuentran los genes, que les protege y permite su multiplicación, pasando de un “envoltorio” a sus descendientes. Esta visión, centrada en los genes, ha sido impulsada por el biólogo inglés Richard Dawkins en su libro The selfish gene (“el gen egoísta”)[17]. Según este enfoque, la selección natural actúa no tanto sobre el organismo, sino también, a nivel más profundo, sobre los genes. El enfoque de Dawkins ha sido criticado de múltiples partes por ser muy reduccionista, ya que no se pueden considerar los genes por separado del organismo en el que residen[18], pero por lo menos llamó la atención sobre la importancia de los genes.
Pero también hay otros niveles sobre los cuales puede actuar la selección natural.
En primer lugar, es posible que la selección natural actúe no sobre genes específicos, sino sobre grupos de genes: tal como existe variación en un gen dentro de una población (las variantes del gen se llaman alelos), puede haber en una especie variantes de grupos de genes[19].
En segundo lugar, es posible que la selección natural actúe no solamente sobre organismos individuales, sino también sobre poblaciones (grupos) de organismos de una misma especie. Esto es lo que se denomina “selección de grupo” (group selection). El planteamiento fundamental de la selección de grupo es que, en grupos de plantas o animales en los que la adecuación de un individuo está vinculado estrechamente al desempeño del grupo, la selección natural favorecerá aquellos rasgos que ayudan a mejorar el éxito del grupo. Este concepto es controvertido, ya que las mutaciones genéticas que dan origen a los cambios de fenotipo por definición se dan en un individuo, para el beneficio del individuo, no del grupo. Pero hay varias consideraciones que apoyan el enfoque de que la selección natural actúa también sobre grupos.
El concepto de la selección de grupo proviene de Darwin. En su libro El origen del hombre, Darwin propone que ocurre una competencia entre grupos de individuos humanos, y que gana aquel grupo cuyos miembros cooperen más y tengan estándares morales más altos[20]. La selección de grupo tiene que ver con altruismo y con acciones realizadas por el bien de grupo, a veces contrarias al interés individual. Por este conflicto con el interés personal, la selección de grupo fue considerada un concepto no válido porque la selección natural actúa por definición en el interés personal, no en contra del mismo. En los años ’60, el gran impulsor de la selección de grupo fue el zoólogo británico Vero Wynne-Edwards, quien planteó que en especies animales hay competencia entre grupos de la misma especie, y que en cada grupo hay algún balance entre los ejemplares cooperadores y los que no lo son[21]. Esta visión de la selección de grupo era un poco ingenua, al pensar – por ejemplo – que un pájaro pudiera elegir no reproducirse por el bien de su grupo. Esta visión fue posteriormente descartada.
También en los años ’60 se propuso la selección de parentesco ó selección familiar (kin selection) como explicación alternativa de la cooperación entre individuos[22]. Se basa en la idea de que si un individuo ayuda a un familiar, que por ser familiar tiene muchos genes en común con el individuo, el individuo favorece la supervivencia de parte de sus propios genes: los que porta el familiar. Esta teoría explica la cooperación entre familiares, por ejemplo en el caso de las colonias de los insectos sociales, donde todos los individuos están relacionados entre sí[23]. También explica una de las observaciones que Wynne-Edwards usó como evidencia para la selección de grupo: que animales limitan sus números para no agotar sus fuentes de alimentos[24]. Sin embargo, la selección familiar no explica la cooperación entre individuos no relacionados, que es relevante por ejemplo en el caso de los humanos. Además, matemáticamente es equivalente a la teoría de selección de grupo, cuando se toma en cuenta la selección tanto al nivel del individuo como al nivel del grupo[25].
Para estudiar la aparición y la permanencia de cooperación entre individuos en un grupo, a veces contraria a los intereses personales de cada individuo, se ha desarrollado en los últimos decenios una rama de ciencia experimental, llamada la teoría del juego (game theory)[26]. Esta estudia, mediante modelos matemáticos y experimentos, qué pasa en un grupo de individuos si no todos cooperan con los demás. Este tema amerita un post separado; limitémonos en este momento a observar que según varios modelos un grupo puede mantener, bajo ciertas condiciones tales como la presencia de una “policía”, un espíritu de cooperación aunque no todos los individuos cooperen[27]. Pero según otros modelos, los cooperantes perderían paulatinamente terreno a los no cooperantes[28].
A pesar de esto, la selección de grupo ha vuelto últimamente a tener cierto auge. Esto se debe especialmente a los esfuerzos del biólogo estadounidense Edward O. Wilson, quien al estudiar colonias de hormigas y otros insectos sociales (en las cuales una mayoría de individuos se sacrifica, quedándose sin descendientes directos, a favor de la colonia en general, y de un individuo, la reina, en particular) llegó a desechar la selección familiar y creó una nueva rama de ciencia, la sociobiología[29]. Wilson y David Sloan Wilson (no un familiar) resumieron la sociobiología así: El egoísmo vence al altruismo dentro de los grupos. Grupos altruistas vencen a grupos egoístas. Todo lo demás es comentario[30]. O sea, los individuos dentro de un grupo compiten entre sí por la supervivencia del mejor adecuado, mediante la selección natural. Pero en la competencia entre grupos, gana aquel grupo cuyos miembros mejor colaboran.
Problemas matemáticos con los modelos de la selección familiar llevaron al biólogo matemático austríaco Martin Nowak a unirse a E.O. Wilson, considerando que la selección familiar no es capaz de explicar la aparición y evolución de las comunidades de los insectos sociales, en una publicación[31] que recibió unos cuantos comentarios negativos de científicos opuestos a la teoría de la selección de grupo[32].
Aunque la sociobiología, basada en los insectos sociales, no es aplicable directamente a las sociedades humanas[33], el concepto de la selección de grupo puede ser utilizado para entender fenómenos humanos tales como los movimientos religiosos, estudiados por David Sloan Wilson[34].
La selección de grupo tiene, en el caso humano, un importante componente cultural; pero aun así, no se puede desligar de la selección natural: si en un grupo humano se produce una mutación genética que predispone los individuos a ser más cooperativos y menos conflictivos, la selección natural causaría un aumento de la ocurrencia de esta mutación en el grupo, que se volvería más cooperativo y, según ya aseveró Darwin, más exitoso comparado con otros grupos humanos con menor grado de cooperación.
En el caso de los demás animales, se ha comprobado recientemente, por primera vez, que la selección de grupo puede determinar ciertos rasgos colectivos de poblaciones. El animal en cuestión es una pequeña araña, Anelosimus studiosus, que vive en colonias elaborando, entre todos los miembros de la colonia, inmensas telarañas – hasta el tamaño de un coche – capaces de atrapar inclusive a pequeños roedores[35]. Las colonias de Anelosimus se caracterizan por una adaptación a nivel de grupo: una relación específica entre hembras agresivas y hembras dóciles. Esta relación depende de la disponibilidad de alimentos y del tamaño de la colonia. Las diferencias en esta proporción estriban en que, aunque por un lado las hembras agresivas son buenas en defender las telarañas de parásitos (que aparecen si hay muchos alimentos), por otro lado tienden a comerse los huevos de sus compañeras – de manera que no es conveniente tener en el grupo más hembras agresivas que lo necesario.
Resulta que la relación dócil/agresiva se mantiene aproximadamente constante de una generación a otra: es un rasgo colectivo, controlado por la selección natural, que determina si una colonia se mantiene o colapsa. En otras palabras, es un caso de selección de grupo. El otro tipo de selección, la selección familiar, no aplica en este caso, ya que las arañas en la colonia no son todas relacionadas entre si.
Así aparece que la evolución, regida por la selección natural (y la selección sexual, como vimos arriba), puede actuar sobre niveles distintos, desde el del gen hasta el del grupo (entendiéndose como grupo cualquier agrupación relevante, que puede variar desde la familia hasta – en el caso de los primates – tribus enteras). Este concepto se llama selección multinivel (multilevel selection).
De hecho, en ciertos respectos se puede considerar a los grupos como organismos. Especialmente en el caso de los insectos sociales, tales como las abejas o las hormigas, sus colonias son como super-organismos: un tipo de organismo compuesto no por células, sino por individuos, y así como las células de un organismo cooperan para la supervivencia del organismo, en el caso del super-organismo son los individuos que componen la colonia los que cooperan para que esta sobreviva[36]. En efecto, hay más paralelos entre los organismos y los super-organismos, por ejemplo en términos de metabolismo, la tasa de crecimiento, o el tiempo de vida: en estos, y otros, aspectos un super-organismo (o sea, una colonia) se comporta de manera parecida a los organismos[37].
Selección sub-óptima
Cabe destacar que la selección natural, aunque resulte en organismos mejor adaptados, no necesariamente su grado de adaptación es el mejor posible. Esto puede tener varias causas. En primer lugar, al nivel genético hay mutaciones que se dan con mayor facilidad que otros, y a menudo ocurre que las mutaciones “fáciles” no resultan en adaptaciones óptimas, sino más bien medianamente buenas[38].
En segundo lugar, al nivel del fenotipo a menudo ocurre que una adaptación buena para una cosa, puede ser mala para otra. El resultado, entonces, suele ser un compromiso entre las dos necesidades. Por ejemplo, el tracto vocal humano está bien adaptado para el habla, pero a costa de un elevado riesgo de problemas dentales (ya que nuestras mandíbulas son demasiado pequeñas para acomodar bien a todos nuestros dientes) y de atragantarnos (ya que nuestras cuerdas vocales ya no sirven para aislar la tráquea – el ducto que lleva aire a los pulmones – cuando estamos comiendo o bebiendo)[39].
Selección natural en otros campos
El concepto de “selección natural” ha sido aplicado también en ámbitos fuera de la biología, por ejemplo en el campo del estudio de la cultura humana, cuya evolución ha sido guiada por procesos muy parecidos a la selección natural (de esto hablaremos en futuros posts en este blog). Sin embargo, es mejor evitar de hablar de selección natural o de survival of the fittest en el caso de los humanos modernos: en el pasado se han utilizado estos conceptos para fines negativos, por ejemplo para justificar la dominación de ciertas razas humanas por otra raza, o la opresión de clases sociales “bajas” por la clase “alta”. Este uso indebido de conceptos científicos afortunadamente ha sido terminado, pero hay que vigilar que no vaya a reaparecer en algún futuro.
Conclusión
La evolución biológica, o sea los cambios en las especies que habitan la faz de la tierra, se dan mediante el mecanismo de la selección natural y mecanismos afines, tales como la selección sexual, la de grupo, la familiar etcétera, que son variaciones de la selección natural. Este mecanismo, postulado en 1859 por Charles Darwin (e, independientemente y casi contemporáneamente, por su coterráneo Alfred Russell Wallace)[40], ha resultado ser la mejor explicación de los cambios en las especies. La teoría ha sido refinada en el tiempo y ha sido llevada al nivel genético, pero su fundación se ha mantenido igual.
Un aspecto importante de la evolución, sin embargo, no se puede explicar suficientemente con el mecanismo de la selección natural: se trata de la aparición de nuevas especies. De este tema hablaremos en el próximo post.
Nota: partes de este post fueron publicadas originalmente en mi blog – ahora cerrado – “Tiempos de Cambio”. La figura en el post fue elaborada por el autor. La foto que acompaña al título es de una jirafa de África occidental (fuente: http://commons.wikimedia.org/wiki/File:Giraffe_koure_niger_2006.jpg).
[1]. Darwin, Ch., 1859/1968. The origin of species by means of natural selection. Penguin Books, Londres.
[2]. Spencer, H., 1864/2002. The principles of biology (2 Vols.). University Press of the Pacific, Honolulu, Hawaii. Ver también: Bowler, P.J., 1983/1989. Evolution: the history of an idea (Ed. rev.). University of California Press, Berkeley, California (pág. 238).
[3]. Wilde, S. y otros, 2014. Direct evidence for positive selection of skin, hair, and eye pigmentation in Europeans during the last 5,000 y. Proceedings National Acedemy of Sciences, 111 (13), 4832-4837. www.pnas.org.
[4]. Ver: http://en.wikipedia.org/wiki/Balancing_selection.
[5]. Durham, W.H., 1991. Coevolution. Genes, culture, and human diversity; pág. 123-153. Stanford University Press. Ver también: http://es.wikipedia.org/wiki/Anemia_falciforme.
[6]. Darwin, 1871/2005. Cáp. VIII-XXI. Ver nota 1.
[7]. Zahavi, A., 1975. Mate selection – A selection for a handicap. Journal of Theoretical Biology, 53, 205-214. http://www.eebweb.arizona.edu/Faculty/Dornhaus/courses/materials/papers/other/Zahavi%20sexual%20selection%20handicap%20model%20signal.pdf. Zahavi, A. y Zahavi, A., 1997. The handicap principle. A missing piece of Darwin’s puzzle. Oxford University Press.
[8]. Zollman, K.J.S., Bergstrom, C.T. y Huttegger, S.M., 2012. Between cheap and costly signals: the evolution of partially honest communication. Proceedings Royal Society B, 280 (1750), 20121878. http://rspb.royalsocietypublishing.org/content/280/1750/20121878.
[9]. Vanpé, C. y otros, 2007. Antler size provides an honest signal of male phenotypic quality in roe deer. The American Naturalist, 169 (4), 481-493. http://www.jstor.org/discover/10.1086/512046.
[10]. Geary, D.C., 1998/2009. Male, female. The evolution of human sex differences. American Psychological Association.
[11]. Knell, R.J., Naish, D., Tomkins, J.L. y Hone, D.W:E., 2012. Sexual selection in prehistoric animals: detection and implications. Trends in Ecology & Evolution, 28 (1), 38-47. http://www.davehone.co.uk/wp-content/uploads/2014/01/Knell-et-al-2012-sexual-selection-in-the-fossil-record.pdf.
[12]. Olsen, B.J., Greenberg, R., Walters, J.R. y Fleischer, R.C., 2013. Sexual dimorphism in a feeding apparatus is driven by mate choice and not niche partitioning. Behavioral Ecology, 24 (6), 1327-1338. http://beheco.oxfordjournals.org/content/24/6/1327.full.pdf+html.
[13]. Bateman, A.J., 1948. Intra-sexual selection in Drosophila. Heredity, 2, 349-368. http://www4.nau.edu/shustercourses/BIO%20698/Literature/Bateman1948.pdf.
[14]. Gowaty, P.A., Kim, Y.-K. y Anderson, W.W., 2012. No evidence of sexual selection in a repetition of Bateman’s classic study of Drosophila melanogaster. Proceedings National Acedemy of Sciences, 109 (29), 11740-11745. www.pnas.org.
[15]. Berglund, J., Pollard, K.S. y Webster, M.T., 2009. Hotspots of biased nucleotide substitutions in human genes. PLoS Biology, 7 (1), e1000026. www.plosbiology.org.
[16]. Williams, T.M., Selegue, J.E., Werner, T., Gompel, N., Kopp, A. y Carroll, S.B., 2008. The regulation and evolution of a genetic switch controlling sexually dimorphic traits in Drosophila. Cell, 134 (4), 610-623. http://www.sciencedirect.com/science/article/pii/S0092867408008404.
[17]. Dawkins, R., 1976/1989. The selfish gene. Oxford University Press.
[18]. Rose, S., 1998. Lifelines. Biology beyond determinism. Oxford University Press.
[19]. Hittinger, C.T., Gonçalves, P., Sampaio, J.P., Dover, J., Johnston, M. y Rokas, A., 2010. Remarkably ancient balanced polymorphisms in a multi-locus gene network. Nature, 464 (7285), 54-58. http://www.nature.com/nature/journal/v464/n7285/full/nature08791.html.
[20]. Darwin, Ch.R., 1871/2005. The descent of man and selection in relation to sex. Elibron Classics, Adamant Media Corp. Cáp. V, pág. 132.
[21]. Wynne-Edwards, V.C., 1962. Animal Dispersion in Relation to Social Behaviour. Oliver & Boyd, London. Ver también: reseña del libro, http://onlinelibrary.wiley.com/doi/10.1111/j.1474-919X.1962.tb08690.x/pdf; comentrio por el mismo autor, http://garfield.library.upenn.edu/classics1980/A1980JV27800001.pdf.
[22]. Hamilton, W.D., 1964. The genetical evolution of social behaviour. I. Journal of Theoretical Biology, 7 (1), 1-16. http://www.sciencedirect.com/science/article/pii/0022519364900396. Hamilton, W.D., 1964. The genetical evolution of social behaviour. II. Journal of Theoretical Biology, 7 (1), 16-52. http://www.sciencedirect.com/science/article/pii/0022519364900396.
[23]. Hughes, W.O.H., Oldroyd, B.P., Beekman, M. y Ratnieks, F.L.W., 2008. Ancestral monogamy shows kin selection is key to the evolution of eusociality. Science, 320 (5880), 1213-1216. http://www.sciencemag.org/content/320/5880/1213.full.pdf.
[24]. Wild, G., Gardner, A. y West, S.A., 2009. Adaptation and the evolution of parasite virulence in a connected world. Nature, 459, 983-986. http://www.nature.com/nature/journal/v459/n7249/full/nature08071.html.
[25]. Wade, M.J. y otros, 2010. Multilevel and kin selection in a connected world. Nature, 463, E8-E9. http://www.nature.com/nature/journal/v463/n7283/full/nature08809.html.
[26]. Ver: http://es.wikipedia.org/wiki/Teor%C3%ADa_de_juegos.
[27]. Por ejemplo: Tomasello, M., Melis, A.P., Tennie, C., Wyman, E. y Herrmann, E., 2012. Two key steps in the evolution of human cooperation. Current Anthropology, 53 (6), 673-692. http://www.jstor.org/stable/10.1086/668207.
[28]. Por ejemplo: Stewart, A.J. y Plotkin, J.B., 2014. Collapse of cooperation in evolving games. Proceedings National Acedemy of Sciences, 110 (38), 15348-15353. www.pnas.org.
[29]. Wilson, E.O., 1975. Sociobiology: The New Synthesis. Harvard University Press.
[30]. Wilson, D.S. y Wilson, E.O., 2007. Rethinking the theoretical foundation of sociobiology. The Quarterly Review of Biology, 82 (4), 327-348. http://www.jstor.org/stable/pdfplus/10.1086/522809.pdf?&acceptTC=true&jpdConfirm=true.
[31]. Nowak, M.A., Tarnita, C.E. y Wilson, E.O., 2010. The evolution of eusociality. Nature, 466, 1057-1062. http://www.nature.com/nature/journal/v466/n7310/abs/nature09205.html. Nowak, M.A., Tarnita, C.E. y Wilson, E.O., 2011. Nowak et al. reply. Nature, 471, E9-E10. http://www.nature.com/nature/journal/v471/n7339/abs/nature09836.html.
[32]. Por ejemplo: Abbot, P. y otros, 2011. Inclusive fitness theory and eusociality. Nature, 471, E1-E4. http://www.nature.com/nature/journal/v471/n7339/full/nature09831.html.
[33]. Wilson, E.O., 2014. The meaning of human existence. Liveright, Norton, New York.
[34]. Wilson, D.S., 2002. Darwin’s cathedral. Evolution, religion, and the nature of society. University of Chicago Press.
[35]. Pruitt, J.N. y Goodnight, C.J., 2014. Site-specific group selection drives locally adapted group compositions. Nature, 514, 359–362. http://www.nature.com/nature/journal/v514/n7522/full/nature13811.html.
[36]. Detrain, C. y Deneubourg, J.-L., 2006. Self-organized structures in a superorganism: do ants “behave” like molecules?. Physics of Life Reviews, 3 (3), 162-187. http://www.sciencedirect.com/science/article/pii/S1571064506000200.
[37]. Hou, C., Kaspari, M., Vander Zanden, H.B. y Gillooly, J.F., 2010. Energetic basis of colonial living in social insects. Proceedings National Acedemy of Sciences, 107 (8), 3634-3638. www.pnas.org.
[38]. Cowperthwaite, M.C., Economo, E.P., Harcombe, W.R., Miller, E.L. y Ancel Meyers, L., 2008. The ascent of the abundant: how mutational networks constrain evolution. PLoS Computational Biology, 4 (7), e1000110. www.ploscompbiol.org.
[39]. Lieberman, Ph., 1992. On the evolution of human language. En: J.A. Hawkins y M. Gell-Mann (editores), The evolution of human languages; 21-47. Santa Fe Institute, Studies in the Science of Complexity, Proceedings Volume XI. Addison-Wesley Publishing Co.
[40]. Ver: http://es.wikipedia.org/wiki/Alfred_Russel_Wallace.

{kind=link}
Saludos Peter, excelente artículo, tome varios apuntes para seguir investigando. Aprovecho la ocasión para tratar de ahondar en un punto que me ha llamado la atención desde hace un tiempo ya: ¿hasta qué punto los seres vivientes son “pasivos” ante el medio? Vale decir; ¿solo pueden adaptarse o morir?
He leído por allí sobre: los “Ingenieros Ecosistémicos” (alogénicos o autogénicos), sobre los organismos “colonizadores” y sobre los primeros organismos “fotosintetizadores”. A propósito de poder “transformar el medio a mi gusto”, teniendo así un rol más “activo” que “pasivo”. No sé si lo tratas más adelante, pues estoy leyendo tus post a destiempo. Disculpa y gracias de antemano.
Me gustaMe gusta
Hola Mauricio, estás tocando un tema muy interesante. Seguro, la vida ha sido un actor importante en la historia de nuestro planeta. La oxigenación de la atmósfera, por ejemplo, es el resultado de – entre otras cosas – la actividad de los organismos aeróbicos (ver la entrada del 18 marzo 2019). La hipótesis de Gaia de Lovelock también le da a la vida un protagonismo en la historia geológica. De esta hipótesis – que por cierto ha sido refutada en buena parte – hablaré en, espero, un futuro próximo.
Me gustaMe gusta