
¿Sabías que las enfermedades – o, mejor dicho, los patógenos que las causan, tales como el virus responsable del COVID-19 – también están sujetas a las leyes de la evolución? ¿Y sabías que los árboles, aunque no se pueden mover, tampoco se escapan a estas leyes? En esta entrada vamos a hablar un poco de la evolución a distintas escalas, y veremos cómo aplican las mismas leyes a pesar de las tallas.
Los patógenos
Los patógenos son los microorganismos – virus, bacterias, etc. – responsables de las enfermedades infecciosas. El COVID-19 es una de éstas, pero también lo son, por ejemplo, la malaria, el dengue, el SIDA, la sífilis, la peste y la tuberculosis. Cuando un patógeno entra en el cuerpo de un huésped (un ser humano, por ejemplo), empieza a multiplicarse y, al mismo tiempo, le causa alguna enfermedad. En efecto, el patógeno necesita al cuerpo del huésped para multiplicarse, y también requiere de los síntomas de la enfermedad para que sus descendientes puedan desplazarse a otro huésped y allí continuar a multiplicarse. La capacidad de un patógeno de causar enfermedad en el huésped se denomina virulencia. En el caso de enfermedades cuyo desenlace puede ser fatal para el huésped, se calcula cuál es el índice de mortalidad de la enfermedad. Patógenos cuya virulencia es muy elevada suelen causar enfermedades con una mortalidad alta [1].
Desde el punto de vista evolutivo, los patógenos se adaptan a sus huéspedes de tal manera que puedan maximizar sus descendientes y el traspaso de éstos a otros huéspedes. En otras palabras: para un patógeno es ideal que pueda multiplicarse libremente en el huésped y que muchos de sus descendientes logren pasar a otros huéspedes, por ejemplo mediante estornudos, tos o excreciones, para repetir el ciclo de la multiplicación y posteriormente el traslado al siguiente huésped. (Esto puede ser bueno para el patógeno, pero obviamente no lo es en absoluto para el desafortunado huésped y los individuos a su alrededor.)
¿Cuál situación es “ideal” para un patógeno? Para el patógeno es bueno causar en su huésped una enfermedad que proporcione suficientes probabilidades de que sus descendientes logren transmitirse a otros huéspedes. O sea: cuantos más estornudos, tos, diarrea o pus cause la enfermedad, mejor. Pero tampoco debe morirse demasiado rápido el huésped, puesto que al morirse éste, también se mueren todos los organismos que lleva adentro, incluyendo al mismo patógeno. Por lo tanto, desde el punto de vista del patógeno existe una virulencia óptima: no tan elevada que el huésped fallezca rápidamente, pero tampoco tan leve que el huésped logre combatir la infección dejando al patógeno sin descendientes [2].
En el caso de enfermedades infecciosas de una virulencia muy alta, tales como la fiebre de Ébola, la mortalidad es tan alta que la situación no es óptima para el patógeno: el huésped se muere rápidamente, y para los servicios médicos es posible aislar el huésped, de manera que el patógeno no puede transmitirse [3]. Es previsible que, en algún momento, se produzca una mutación del virus del Ébola que tenga una virulencia menor, con mejores probabilidades de transmisión – por lo menos, es así que actúan los mecanismos evolutivos. En el caso de la sífilis [4], algo parecido parece haber pasado: cuando apareció, al final de siglo XV, tenía una mortalidad muy alta, que rápidamente se redujo, de manera que la enfermedad pudo transmitirse con más éxito.
El COVID-19, por otro lado, es una enfermedad cuya virulencia, desde el punto de vista del patógeno, el coronavirus SARS-CoV-2, no es tan extrema: por lo general, el huésped sobrevive a la infección y la transmisión ocurre fácilmente, de manera que los descendientes del patógeno pueden pasar a una multitud de otros huéspedes y así asegurar al patógeno que la especie no sólo se mantenga en el tiempo, sino también pueda mantener una expansión significativa. Aun así, la virulencia del SARS-CoV-2 parece atenuarse [5], de manera que posiblemente vaya a tener más tiempo para la transmisión a otros huéspedes. Mutaciones del virus pueden resultar en la aparición de variantes con una virulencia aun más cercana a la óptima – o, simplemente, capaces de resistir a los medicamentos que se están desarrollando, de manera que el virus pueda continuar sus ciclos de infección y transmisión.
Esto ya nos muestra que los patógenos son capaces de mutar fácilmente, o sea, su evolución es rápida y ocurre en tiempo real. Los patógenos actuales que nos pueden afectar, están todos sujetos a cambios evolutivos, y en muchos casos, es la presión ejercida por los humanos (por ejemplo, el desarrollo de un antibiótico nuevo) lo que causa, a nuestro pesar, la evolución de los patógenos.
En términos generales, existe una coevolución entre los patógenos y sus huéspedes. Una especie que es afectada por una enfermedad infecciosas, puede – a la larga – desarrollar defensas contra ésta; si estas defensas son heredables, se considera un caso de (micro)evolución. Sin embargo, cuando llegue este momento, el patógeno causante de la enfermedad puede mutarse y, mediante un cambio evolutivo, cambiar a una nueva forma, contra la cual su huésped todavía no tiene resistencia. Este proceso de cambios recíprocos en un patógeno y su desafortunado huésped, es un ejemplo de coevolución antagónica, en el que dos especies, normalmente un depredador y su presa, cambian en respuesta a cambios en su adversario. En la naturaleza se conocen varios casos probables de tal coevolución. En un estudio experimental de pinzones norteamericanos y el patógeno bacterial Mycoplasma gallisepticum se pudo estudiar el proceso en detalle y confirmar esta interacción entre un patógeno y su huésped [6].
En estudios experimentales sucesivos se pudo determinar que, en efecto, una virulencia intermedia es óptima para un patógeno, y que los mismos procesos evolutivos (denominados evolución estabilizadora) conducen a tal condición. Esto contradice la idea de que la virulencia de un patógeno se reduce en el tiempo hasta llegar a niveles mínimos [7].

Fermento
Fermento, o levadura, es el nombre que se da a ciertos hongos microscópicos (recordar que los hongos forman su propio reino, separado de los reinos de los animales y las plantas). Muchos fermentos son beneficiosos, y en efecto, algunos han sido domesticados por los humanos, entre otras cosas para elaborar pan, cerveza, o queso (ver la entrada de 7 marzo 2020). Su utilidad se extiende a los estudios genéticos, incluyendo al estudio de los procesos evolutivos. Al ser posible estudiar sus cambios en el laboratorio, en tiempo real y bajo condiciones controladas por los humanos, esto los convierte en unos de los organismos predilectos de los investigadores.
Uno de los procesos genéticos responsables de los cambios evolutivos de un organismo, es la duplicación de genes. Normalmente los organismos tienen una sola copia de cada gen, pero en algunos casos, poco comunes, un organismo puede quedarse con dos copias de un mismo gen, como resultado de un error en la replicación. Al tener un organismo dos genes que básicamente cumplen la misma función, si ocurre una mutación en uno de los dos, el organismo sigue funcionando igual de bien, al mismo tiempo que el gen mutado puede, dependiendo de la mutación, asumir un papel nuevo que potencialmente puede causar cambios evolutivos [8].
Los genes duplicados se pueden estudiar bien en el fermento Saccharomyces cerevisiae, que es el que se usa para la fermentación alcohólica, pero una cepa del cual se utiliza como levadura de pan (ver imagen). El proceso de duplicación no ha sido observado (ocurrió hace millones de años en el caso de este fermento), pero sí se ha podido estudiar qué puede pasar con los genes duplicados. Los duplicados de ciertos genes no pueden mutarse sin perder efectividad, así que estos duplicados se conservan en el tiempo o se convierten en “basura”. Pero otros genes sí pueden experimentar mutaciones que resultan en una función nueva del gen duplicado: por ejemplo, puede causar la elaboración de una nueva proteína que le confiere al organismo una nueva propiedad, beneficiosa. En este caso, el gen duplicado empieza a tener su propia función y con el tiempo, después de muchas generaciones y unas cuantas mutaciones, ya será muy diferente del gen original [9].
Otro de los mecanismos que pueden causar modificaciones heredables en organismos, identificado hace pocos años, es el de cambios epigenéticos. La epigenética es el estudio de los mecanismos que regulan, mediante marcadores adheridos al ADN, la expresión de los genes, o sea, que definen cuáles genes se leen (expresan), y cuáles no [10]. Estos marcadores pueden ser colocados, movidos o quitados durante la vida del organismo (por ejemplo en función de cambios ambientales), causando cambios en la expresión de los genes, y recientemente se descubrió que estos cambios son heredables. O sea, se trata de microevolución causada por cambios en la expresión de los genes, en lugar de cambios en el material genético mismo. En otras palabras: la evolución no sólo sigue los principios de Darwin, sino a veces los de Lamarck, quien postuló que cambios experimentados por un organismo durante su vida pueden ser heredados por sus descendientes.
Volviendo a los fermentos, esos organismos cuya evolución puede ser estudiada en tiempo real en el laboratorio: los fermentos han ayudado a entender cómo funciona la selección natural mediante los cambios epigenéticos. En un cultivo de fermentos, en el laboratorio, se seleccionaron aquellos organismos cuyo gen regulador de la galactosa (una biomolécula relacionada a los azúcares) tenía una expresión mínima, por debajo de lo normal, debido a la acción de un mecanismo epigenético. Se determinó que los descendientes de estos fermentos mantuvieron, durante varios días (o sea, decenas de generaciones, ya que estos fermentos se multiplican cada cien minutos), esta regulación anómala del gen de la galactosa. Así se confirmó que los cambios epigenéticos son heredables y pueden afectar la adaptación de múltiples generaciones de descendientes [11].

Animales pequeños, grandes lecciones
Pasando de los hongos a los animales, nos detendremos en primer lugar en las mariposas. Las mariposas no son inmunes a la evolución contemporánea: en efecto, en la entrada de 31 julio 2020 ya hablamos de una polilla de las islas británicas que cambió dos veces de color: primero de claro a oscuro, al empezar la Revolución Industrial, cuando aumentó la contaminación; y después de vuelta a claro, cuando bajó la contaminación.
Pero también hay otros ejemplos, entre los que se encuentra una de las mariposas más famosas: la monarca (ver imagen). La mariposa monarca vive principalmente en Norteamérica, donde es famosa por su larga migración anual: desde Canadá a México, y de vuelta [12]. Pero también existen colonias de mariposas monarca, que no migran, en el Caribe, en Suramérica, y en ciertas islas del Pacífico y el Atlántico; estas colonias son descendientes de ejemplares que durante la migración anual fueron desviados de su ruta, por ejemplo por tempestades. Las monarcas en estas colonias, que no migran, tienen las alas más pequeñas que las de las monarcas de las que descendieron, que sí migran. Este cambio, que se dio dentro de los últimos dos siglos, refleja cómo un rasgo favorecido por la necesidad de migrar, se convierte en desfavorecido cuando la población deja de migrar, con el consecuente cambio evolutivo de alas grandes a alas más pequeñas [13].
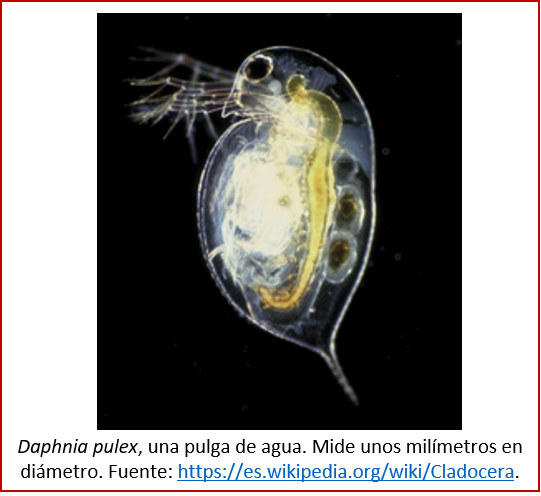
Son muchos los animales pequeños que nos enseñan cómo funciona la evolución. Las pulgas de agua de la especie Daphnia, por ejemplo, que a pesar de su nombre informal son pequeños crustáceos de agua dulce (ver imagen abajo), fueron el objeto de un estudio reciente acerca de la evolución en acción. En lagos del Wisconsin (Estados Unidos), las Daphnia han tenido que encarar la aparición de un nuevo depredador (otra especie de pulga de agua) en el pasado reciente; lo hicieron mediante cambios en conducta y en su historia de vida. Se tomaron muestras de sedimento en el fondo de algunos de estos lagos, con edades distintas: correspondientes a intervalos justo antes, durante, y después de la aparición del depredador. En las muestras había huevos de Daphnia, que pudieron ser resucitados, dando origen a ejemplares vivos, pero de distintos momentos en el tiempo. Estos ejemplares se estudiaron genéticamente.
Se pudo determinar que, antes de la aparición del nuevo depredador, había una variabilidad genética importante. Esto permitió que, cuando hubo que adaptarse al recién llegado depredador, la selección natural pudo actuar eficazmente, al haber variantes genéticas que condujeron a ejemplares de Daphnia mejor adaptados. Pero, aunque unas variantes resultaron ser las más exitosas en términos de adaptación, no por ello desaparecieron las otras variantes: más bien, se mantuvo una variabilidad genética importante. De esta manera, Daphnia estará lista para hacer frente a otros cambios en el entorno, cuando se produzcan [14].
Interacciones
Muchos procesos evolutivos son causados por interacciones con otros organismos – ya lo vimos arriba, en el caso de los patógenos. Una especie está sometida a fuerzas que le empujan a cambiar cuando cambian otras especies en su entorno, de las cuales de una manera u otra depende. Conocido es, en este sentido, lo que se llama la carrera armamentista evolutiva: una especie que tiene depredadores cambia para reducir la probabilidad de que acabe como la cena de un depredador, y el depredador a su vez cambia para que su presa no se le escape. No es coincidencia, por ejemplo, que los herbívoros más rápidos – las gacelas – viven en zonas donde existen los depredadores más veloces. Esta una evolución sin fin, que empezó hace millones de años, que sigue en nuestros tiempos y que continuará. Veamos algunos ejemplos más.
En ciertos casos, los depredadores no son carnívoros, sino otras clases de animales – tales como los animales que se alimentan de productos vegetales. Las ardillas, por ejemplo, mantienen una competencia con los árboles cuyas semillas comen. Para minimizar las pérdidas de semillas debido a la actividad de las ardillas, ciertos árboles, tanto en Norteamérica como en Europa, desarrollaron una estrategia interesante: varían la cantidad de semillas que producen, de manera que en ciertos años producen muchas, y en otros, muy pocas; además, de una manera u otra, los árboles en una región sincronizan estos ciclos, así que producen muchas semillas, o pocas, todos a la vez. El impacto sobre las poblaciones de ardillas (y otros animales que se alimentan de las semillas) es grande: como consecuencia de los años de poca producción de semillas, las poblaciones de ardillas se mantienen limitadas, de modo que cuando hay mucha producción de semillas, no hay suficientes ardillas para que todas las semillas sean comidas. Pero, las ardillas están contraatacando: logran captar, de una manera u otra, los señales de que los árboles se están preparando para producir muchas semillas, y justo antes de ese momento producen una camada de crías adicional a la normal: de esta manera aumenta la población de ardillas justo a tiempo para la bonanza de semillas [15]. Así que, por los momentos, parecen haber ganado las ardillas. Pero en algún momento serán los árboles los que pensarán en una nueva estrategia.
En otros casos, la competencia ocurre entre herbívoros que se alimentan de la misma fuente. Distintas especies de herbívoros grandes, por ejemplo, tienden a nutrirse del mismo tipo de alimentos. Lo que ocurre en tales casos, es una división en nichos: herbívoros distintos suelen nutrirse de diferentes plantas, o de diferentes partes. Algo parecido ha ocurrido en el caso de las jirafas: con su cuello largo, logran alcanzar los topes de los árboles, que los demás herbívoros no alcanzan, de manera que tienen allí su nicho alimenticio [16].
Una instancia interesante de competencia se da entre ciertas especies de plantas, al defenderse ante los insectos herbívoros que comen sus hojas y otras partes del organismo. Las plantas, al no poder huir de sus depredadores herbívoros, desarrollan distintos mecanismos de defensa, tales como espinas, la producción de substancias químicas desagradables, o el desarrollo de hojas tan duras que es difícil para los insectos comerlas. Durante mucho tiempo los investigadores pensaban que los insectos reaccionaban a la aparición de nuevas defensas adaptándose a las mismas, desarrollando ellos mismos nuevos mecanismos para contraatacar a las plantas. Sin embargo, no siempre la evolución funciona así: por ejemplo, se ha detectado en la Amazonia una situación distinta, en la que, cuando una especie vegetal (en este caso específico, un árbol del género Inga) desarrolló defensas nuevas, los insectos depredadores se mudaban a otras especies en lugar de desarrollar nuevas estrategias para seguir consumiendo las hojas de Inga. De esta manera, parece que, por lo menos en algunos casos y por algún tiempo, las especies vegetales logran ganarla a sus depredadores, y que la competencia se da más bien entre distintas especies de vegetales, para ver cuál es la primera en desarrollar defensas efectivas [17].
Otro ejemplo de esto fue descrito para álamos de Norteamérica. Dentro de una misma especie de estos árboles, unos ejemplares tenían mayor concentración de compuestos químicos anti-insectos, de manera que sufrían menos que sus congéneres los ataques a sus hojas de herbívoros tales como polillas y orugas [18].

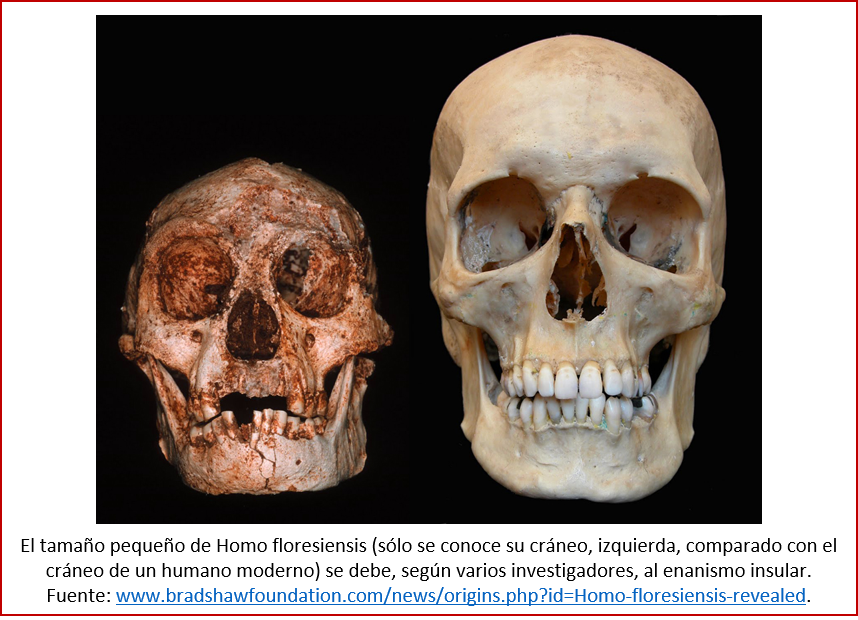
Volviendo a los mamíferos, un caso distinto se ha dado en ciertas islas, por ejemplo en el Mediterráneo, donde se ha observado una reducción del tamaño de varios grupos de mamíferos, lo que se denomina el “enanismo insular”. Hasta hace unos 4500 años, hubo elefantes (ver imagen) y otros mamíferos miniaturizados en la isla de Sicilia. En otras islas se encontraron los restos de pequeños hipopótamos y caballos. Este fenómeno (algunos lo llaman una regla, pero tiene unas cuantas excepciones) se dio, en el caso de los herbívoros medianos y grandes, en islas donde había poca presión de depredadores y poca competencia por los alimentos. También entre los antecesores de nuestra especie puede haber habido casos de enanismo: Homo floresiensis, un congénere nuestro miniaturizado de la isla de Flores (Indonesia; ver imagen abajo), puede deber su tamaño reducido a esta misma causa [19].
Entre los depredadores, el enanismo no ha sido tan marcado, pero en los casos que ocurrió, fue al parecer por la reducción del tamaño de sus presas y/o por una disminución en la cantidad de presas – en tales casos, no tiene sentido que los depredadores se mantengan grandes [20]. El estudio de zorros enanos en islas frente a la costa de California mostró que el proceso del enanismo no sólo implica una reducción de talla, sino también cambios en las formas de las extremidades y otras partes del cuerpo [21]. En otros grupos de animales, sin embargo, no se observó este enanismo; más bien, algunos grupos, por ejemplo ciertos roedores, aumentaron en tamaño [22].
De esta manera, las interacciones entre especies nos muestran que la evolución en acción juega un papel nada desdeñable dentro de los ecosistemas: los cambios evolutivos pueden ocurrir a escalas temporales y geográficas suficientemente finas como para causar cambios que afectan la manera en la que interactúan ciertas especies [23].
Hibridación
Volviendo nuestra mirada a los procesos genéticos que causan la aparición de especies nuevas, vale la pena considerar un proceso típico (mas no exclusivo) de las plantas: se trata de la hibridación. Ya se habló del tema en la entrada de 31 julio 2020. La hibridación consiste en el apareamiento exitoso de dos ejemplares de especies distintas. Esto se da especialmente en el caso del reino vegetal, donde es común la duplicación del material genético.
La hibridación es un proceso que se puede estudiar prácticamente en tiempo real. Dentro de los últimos 150 años apareció en las Alpes suizos una pequeña mata nueva, Cardamine insueta (ver imagen abajo), una especie de berro amargo. Se desarrolló en los últimos 150 años debido a cambios ambientales en el valle circundante, cuando la gente local despejó el bosque y lo convirtió en tierra de pasto. C. insueta se formó a partir de dos especies parentales con hábitats ecológicos específicos: mientras que C. amara crece junto a corrientes de agua, C. rivularis habita en sitios menos húmedos. La conversión del uso de la tierra de bosque a pastizal indujo la hibridación de los dos progenitores generando la nueva especie que se encuentra entre los hábitats de los padres con fluctuación temporal del nivel del agua. Es la combinación de rasgos genéticos de sus padres lo que permitió que la nueva especie creciera en un nicho ambiental distinto [24].

También los árboles son susceptibles a hibridación. Los castaños, por ejemplo, que para fertilizar sus flores necesita de polen de otro castaño, se mezclan fácilmente, especialmente si florecen en las mismas épocas (ver imagen). Se ha podido determinar que una variante china, Castanea henryi var. Omeiensis, es el producto de la hibridación de dos otras especies de castaño que viven en la misma región. Dado que se da tanta mezcla entre castaños, la aparición de una nueva variante estable, que además está encaminada a convertirse en una especie separada, no es sencilla. Sin embargo, resulta factible por la aparición, a nivel genético, de cambios en ciertos cromosomas que forman barreras para la mezcla. Estas barreras le proporcionan a la nueva variante una estabilidad genética, que además parece tener ventajas competitivas; esto hizo que la nueva variante se mantuviera en el tiempo [25].

La marcha de los robles
Puede parecer raro, pero los árboles no sólo están expuestos a los vaivenes de los procesos de la evolución; además, tienden a migrar. Obviamente esta migración no se da al nivel de ejemplares individuales – una de las características de las plantas es que en principio no se mueven –, sino al nivel de la especie. Los descendientes de un árbol crecen a cierta distancia de su progenitor, de manera que, a lo largo de múltiples generaciones, el área de distribución de la especie cambia; cambios climáticos, por ejemplo, pueden resultar en un desplazamiento de esta área hacia el sur o el norte.
Los robles (ver la portada de esta entrada) se desarrollaron en los bosques boreales, en algún momento en el Paleoceno, después de la extinción de los dinosaurios. Eran tiempos cálidos, y a la medida en que las temperaturas bajaban, los robles empezaron a migrar hacia el sur, con ramificaciones hacia Norteamérica y Eurasia. Hoy en día existen ocho linajes de robles; dentro de cada linaje, las distintas especies de robles se mezclan libremente, tal como en el caso de los castaños, dando origen a híbridos con características de ambos progenitores. Esta hibridación ha ayudado a que los robles se adaptaran a distintos entornos: desde ambientes húmedos hasta entornos secos, con ocurrencia de incendios. En cada ambiente, los robles se adaptaron a la situación específica. De esta misma manera, los robles pudieron reaccionar rápidamente cuando, hace veinte mil años, terminaron las glaciaciones, y se tiene la esperanza de que, con el cambio climático actual, los robles serán capaces de ajustarse con la misma efectividad [26].
Conclusión
Esta entrada muestra cómo los procesos de la evolución actúan sobre todos los organismos, desde los más pequeños a los más grandes; animales, vegetales, bacterias, virus, hongos; los lentos, los rápidos…
Muestra también que la naturaleza sigue estando “en obras”: no se puede hablar de una situación específica (por ejemplo, la actual) como fin de la evolución. Más bien, la evolución es un proceso de nunca acabar. Hemos visto que está continuando en nuestros días, a veces lentamente, a veces en un abrir y cerrar de ojos. Generalmente, el proceso es impulsado por cambios en el entorno de origen natural; pero otras veces estos cambios son obra de nosotros, los humanos: bien sea dentro de los muros de un laboratorio, bien sea afuera, a raíz del efecto que tiene nuestra presencia sobre el planeta Tierra.
Del efecto que tenemos los humanos sobre la evolución de la vida terrestre, volveremos a hablar en la próxima entrada.
Nota: la foto en el encabezado del post muestra un antiguo roble. Crédito: Frederika Rademaker. Fuente: https://nl.paganfederation.org/files/2013/12/Oak-Trees.jpg.
[1] Ver, por ejemplo: https://es.wikipedia.org/wiki/Agente_biol%C3%B3gico_pat%C3%B3geno; https://es.wikipedia.org/wiki/Virulencia.
[2] Price, M., 2007. Pathogen virulence: the evolution of sickness. Science Creative Quarterly. www.scq.ubc.ca/pathogen-virulence-the-evolution-of-sickness.
[3] Ver, por ejemplo: https://es.wikipedia.org/wiki/Enfermedad_por_el_virus_del_%C3%89bola.
[4] LaFond, R.E. y Lukehart, S.A., 2006. Biological basis for syphilis. Clinical Microbiology Reviews, 19 (1), 29-49. https://doi.org/10.1128/CMR.19.1.29-49.2006.
[5] Lan, F.-Y., Filler, R., Mathew, S., Iliaki, E., Osgood, R., Bruno-Murtha, L.A. y Kales, S.N., 2020. Evolving virulence? Decreasing COVID-19 complications among Massachusetts healthcare workers: a cohort study. medRxiv. https://doi.org/10.1101/2020.08.17.20176636.
[6] Bonneaud, C., Giraudeau, M., Tardy, L., Staley, M., Hill, G.E. y McGraw, K.J., 2018. Rapid antagonistic coevolution in an emerging pathogen and its vertebrate host. Current Biology, 28, 1-6. https://doi.org/10.1016/j.cub.2018.07.003.
[7] Bonneaud, C., Tardy, L., Hill, G.E., McGraw, K.J., Wilson, A.J. y Giraudeau, M., 2020. Experimental evidence for stabilizing selection on virulence in a bacterial pathogen. Evolution Letters, publicado online. https://doi.org/10.1002/evl3.203.
[8] Ver: https://es.wikipedia.org/wiki/Duplicaci%C3%B3n_cromos%C3%B3mica.
[9] Kuzmin, E. y otros, 2020. Exploring whole-genome duplicate gene retention with complex genetic interaction analysis. Science, 368 (6498), eaaz5667. https://doi.org/10.1126/science.aaz5667.
[10] Ver: https://es.wikipedia.org/wiki/Epigen%C3%A9tica.
[11] Luo, X., Song, R., Moreno, D.F., Ryu, H.-Y., Hochstrasser, M. y Acar, M., 2020. Epigenetic mechanisms contribute to evolutionary adaptation of gene network activity under environmental selection. Cell Reports, 33, 108306. https://doi.org/10.1016/j.celrep.2020.108306.
[12] Ver: https://es.wikipedia.org/wiki/Danaus_plexippus.
[13] Freedman, M.G., Dingle, H., Strauss, S.Y. y Ramírez, S.R., 2020. Two centuries of monarch butterfly collections reveal contrasting effects of range expansion and migration loss on wing traits. Proceedings of the National Academy of Sciences, 117 (46), 28887-28893. https://doi.org/10.1073/pnas.2001283117.
[14] Landy, J.A., Oschmann, A., Munch, S.A. y Walsh, M.R., 2020. Ancestral genetic variation in phenotypic plasticity underlies rapid evolutionary changes in resurrected populations of waterfleas. Proceedings of the National Academy of Sciences. https://doi.org/10.1073/pnas.2006581117.
[15] Boutin, S., Wauters, L.A., McAdam, A.G., Humphries, M.M., Tosi, G. y Dhondt, A.A., 2006. Anticipatory reproduction and population growth in seed predators. Science, 314 (5807), 1928-1930. https://doi.org/10.1126/science.1135520.
[16] Cameron, E.Z. y du Toit, J., 2007. Winning by a neck: tall giraffes avoid competing with shorter browsers. The American Naturalist, 169 (1), 130-135. https://doi.org/10.1086/509940.
[17] Endara, M.-J. y otros, 2017. Coevolutionary arms race versus host defense chase in a tropical herbivore–plant system. Proceedings of the National Academy of Sciences, 114 (36), E7499-E7505. https://doi.org/10.1073/pnas.1707727114.
[18] Hemming, J.D.C. y Lindroth, R.L., 1995. Intraspecific variation in aspen phytochemistry: effects on performance of gypsy moths and forest tent caterpillars. Oecologia, 103, 79-88. https://doi.org/10.1007/BF00328428.
[19] Kubo, D., Kono, R.T. y Kaifu, Y., 2013. Brain size of Homo floresiensis and its evolutionary implications. Proceedings of the Royal Society B, 28020130338. http://doi.org/10.1098/rspb.2013.0338. Ver también: www.sciencemag.org/news/2013/04/scienceshot-shrinking-hobbits-brain.
[20] Raia, P. y Meiri, S., 2006. The island rule in large mammals: paleontology meets ecology. Evolution, 60 (8), 1731-1742. https://doi.org/10.1111/j.0014-3820.2006.tb00516.x.
[21] Young, C.B., 2020. Static allometry of a small-bodied omnivore: body size and limb scaling of an island fox and inferences for Homo floresiensis. Journal of Human Evolution, 149. https://doi.org/10.1016/j.jhevol.2020.102899.
[22] Meiri, S., Cooper, N. y Purvis, A., 2008. The island rule: made to be broken? Proceedings of the Royal Society B, 275, 141-148. http://doi.org/10.1098/rspb.2007.1056.
[23] Urban, M.C. y otros, 2020. Evolutionary origins for ecological patterns in space. Proceedings of the National Academy of Sciences, 117 (30), 17482-17490. https://doi.org/10.1073/pnas.1918960117.
[24] Sun, J., Shimizu-Inatsugi, R., Hofhuis, H., Shimizu, K., Hay, A., Shimizu, K.K. y Sese, J., 2020. A recently formed triploid Cardamine insueta inherits leaf vivipary and submergence tolerance traits of parents. Frontiers in Genetics, 11, 567262. https://doi.org/10.3389/fgene.2020.567262. Ver también: https://mundoagropecuario.com/evolucion-en-accion-una-nueva-especie-vegetal-en-los-alpes-suizos.
[25] Sun, Y., Lu, Z., Zhu, X. Y Ma, H., 2020. Genomic basis of homoploid hybrid speciation within chestnut trees. Nature Communications, 11, 3375. https://doi.org/10.1038/s41467-020-17111-w.
[26] Hipp, A.L., Manos, P.S. y Cavender-Bares, J., 2020. Ascent of the oaks. Scientific American, agosto 2020, 42-49. www.scientificamerican.com/article/how-oak-trees-evolved-to-rule-the-forests-of-the-northern-hemisphere.

{kind=link}
2 comentarios en “Evolución contemporánea (5): un cuento de nunca acabar”