
La raza humana es, sin lugar a dudas, una de las especies animales más exitosas que habiten la tierra, y la más poderosa en términos de su capacidad de alterar a su entorno. Hoy en día, mucha gente está preocupada – y con razón – por el impacto negativo que puede tener nuestra especie sobre nuestro planeta, alterando el clima, afectando a los ecosistemas, contaminando las aguas y el aire… Sin embargo, la medalla tiene dos caras: si somos los más poderosos a la hora de alterar nuestro entorno, también somos la especie que, después de nacer en humilde cuna, hemos logrado un desarrollo que, mediante cambios genéticos y culturales, nos permitió convertirnos en la especie exitosa que ahora somos.
Con el presente post voy a empezar una serie de varias entregas en las cuales consideraré algunos de los eventos y procesos que condujeron al éxito humano. Empezaremos por el origen de nuestra especie.
Nuestra especie: Homo sapiens
En el reino animal, al cual pertenecemos, así como en los demás reinos de la naturaleza, todas las especies conocidas han recibido un nombre oficial y han sido agrupadas según sus características. El padre de este sistema fue el naturalista sueco Carlos Linneo (1707-1778), quien le puso nombres científicos a las especies que conocía, y las agrupó según sus características: las especies las agrupó en géneros, los géneros en familias, las familias en órdenes, los órdenes en clases, y las clases en reinos[1]. Todas las especies llevan un nombre que consiste de dos partes: la primera, indicando el género al que una especie pertenece, y la segunda, el nombre de la especie. El nombre científico del león, por ejemplo, es Panthera leo, Panthera siendo el nombre del género del que el león forma parte. Hoy día se sigue utilizando esta clasificación taxonómica de Linneo, con algunas modificaciones.
Considerando las similitudes entre el hombre y los simios, Linneo se dio cuenta de que taxonómicamente ambos forman parte de un mismo orden, al que llamó los primates. En círculos religiosos no fue muy bien recibida esta clasificación del hombre junto con los simios, pero de punto de vista científico resultó ser la correcta. Pero los humanos sí fuimos asignados a un género propio, Homo, y el nombre científico que Linneo nos dio, es Homo sapiens, el “hombre sabio”.
Así que nosotros formamos parte de la especie Homo sapiens (que podemos abreviar como H. sapiens). Se ha intentado subdividir nuestra especie en razas, y de hecho hay cierta discusión acerca de diferencias fisiológicas y genéticas entre distintas poblaciones humanas, pero el mero hecho de que no existe acuerdo sobre la definición de las razas humanas, ni de cuántas hay (los números varían de tres a cien), demuestra que no tiene mucho sentido hacer énfasis en el concepto de las razas: todos los humanos pertenecemos a la misma especie[2].
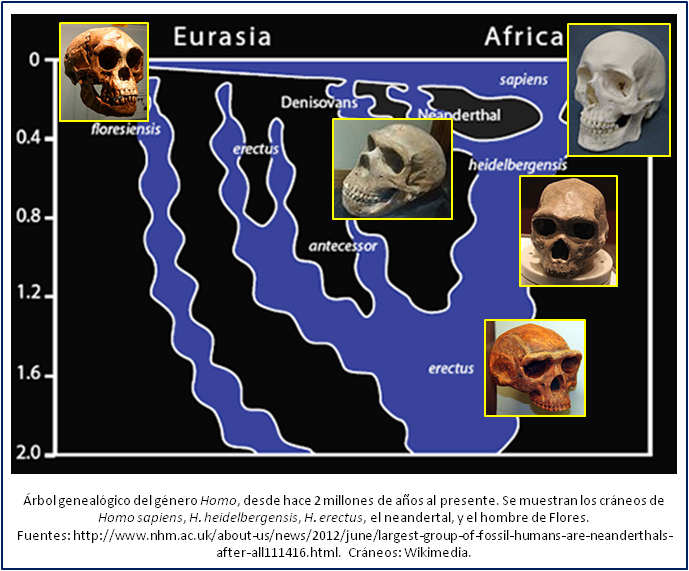
Nuestros primos
En la actualidad somos nosotros los únicos representantes del género Homo. Sin embargo, hace poco tiempo, sólo unas decenas de miles de años, teníamos varios congéneros. Llamémoslos nuestros “primos”. Los más famosos son los neandertales. Pero hubo varios más, de los cuales hoy día conocemos al hombre de Denisova y al de Flores.
El hombre de Neandertal, H. neanderthalensis, es el primer ancestro de nuestra especie cuyos restos se encontraron – en Alemania y Bélgica, en el siglo XIX. En la figura que acompaña este post se puede apreciar la imagen de un cráneo de un neandertal. A diferencia de nosotros tiene un arco supraciliar (una protuberancia por encima de los ojos) marcado, y su frente es menos vertical que la nuestra. También tenía una nariz un poco más grande. Pero del resto, su aparencia y estatura no fueron muy distintas de las nuestras. Los restos más viejos que sin duda pertenecen a un neandertal, encontrados en Inglatierra, datan de hace 400.000 años[3], mientras que los últimos neandertales pueden haber vivido en el sur de la península iberica, hace unos 30.000 años[4]. Hasta la fecha no se sabe con seguridad qué causó la desaparición de los neandertales, aunque abundan las teorías (en un post futuro hablaré de éstas). Se ha logrado obtener restos del ADN, es decir, el material genético, del varios neandertales, lo que permitió leer buena parte de su genoma y compararlo con el nuestro[5]; abajo volveremos a hablar de esto.
Del hombre de Denisova se conoce sólo un hueso de un dedo, hallado en una gruta en el sur de Siberia, así que no sabemos mucho de él. El dedo perteneció a una niña que vivió hace unos 30.000 años. Con tan poco material no es posible darle un nombre oficial; pero se pudo leer su ADN, lo que permitió determinar que era genéticamente distinto a nosotros y también al neandertal, su “primo” más cercano[6]. En España se encontraron, en el material genético de un individuo (un neandertal, o miembro de una especie más vieja, Homo antecessor) que vivió hace 400.000 años, secuencias de ADN típicas del hombre de Denisova; cómo hayan logrado estas secuencias llegar hasta allá es una incógnita que aun queda por aclarar[7].
El tercero de nuestros primos es el hombre de Flores. Flores es una isla de Indonesia, donde, en una cueva, se encontró un cráneo muy parecido al humano, pero mucho más pequeño, perteneciente a un individuo que vivió hace unos 18.000 años[8]. Este hallazgo causó una fuerte polémica en el mundo científico: algunos aseveraron que se trata de un individuo con microcefalia, o sea una malformación del cráneo, mientras que según otros se trata de una especie nueva, H. floresiensis[9]. Los últimos parecen tener los argumentos más válidos, por el momento. Desafortunadamente no se pudo recuperar el ADN del hombre de Flores, ya que no quedó conservado debido al clima tropical.
En búsqueda de nuestro ancestro
Nuestra especie, H. sapiens, no apareció de la nada, y nuestros primos, desde luego, tampoco. Tal como ya observó Linneo, tenemos mucho en común con los simios, y es lógico suponer que formamos parte de un mismo linaje antiguo, que en algún momento se separó en dos: uno, conduciendo a los simios actuales, y otro, pasando por el género Australopithecus, al género Homo – primero los Homo arcaicos, y al final nosotros. En otra oportunidad escribiré acerca de la separación de los dos linajes, y de los primeros Homo. Por los momentos deseo considerar únicamente a nuestro ancestro directo.
Entonces, ¿cuál es este ancestro? Por extraño que eso pueda parecer, todavía no lo sabemos con seguridad. Afortunadamente los estudios genéticos nos han permitido aclarar algunos asuntos.
Una gran duda que lograron despejar los estudios genéticos, es si H. sapiens se originó en un solo lugar, África, esparciéndose desde allí por el mundo, o si se desarrolló por separado en distintos lugares, es decir no sólo en Africa, sino también en la China y otros sitios[10]. Esta última hipótesis, llamada “multirregional”, fue rechazada a favor de la primera, la que postula un origen en África[11]. Desde los años ’80, cuando empezó a ser posible examinar en primer lugar el material genético de las personas vivas, y luego también el de los restos humanos fósiles, quedó claro que H. sapiens surgió en África hace unos 200.000 años. Se determinó primero que todos nuestros antepasados vienen de un solo lugar, en el África subsahariana[12]. Investigaciones genéticas recientes indicaron que el grupo de población que está más cerca del primero H. sapiens, es el de los bosquimanos. Éstos se encuentran hoy en día sólo en el sur de África, pero originalmente vivían también en otras partes de ese continente[13].
Los restos fósiles confirman nuestro origen africano. En 2003 se dio a conocer el hallazgo, hecho en Etiopía, de algunos cráneos de individuos de H. sapiens, con una edad de 160.000 años (aunque no lucen exactamente como nosotros y por lo tanto se considera que se trata de una subespecie distinta, a la que se dio el nombre de H. sapiens idaltu, mientras que nosotros seríamos H. sapiens sapiens)[14]. Dos años después se determinó que algunos cráneos del H. sapiens moderno (o sea, parecido a nosotros), encontrados cerca del río Omo, también en Etiopía, son aun más viejos, teniendo una edad de 195.000 años[15]. Esta edad coincide muy bien con la de 200.000 años, obtenida por los estudios genéticos, para nuestra especie.
Así que, al tener un solo origen, tenemos un sólo ancestro. Los estudios genéticos nos permiten decir algo acerca de cuándo vivió el último ancestro común de nuestra especie y de los neandertales – aunque no nos dice quién fue. Al comparar el ADNmt, o sea el ADN de las mitocondrias (orgánulos celulares que tienen su propio material genético) de las dos especies, se ha determinado que el último ancestro común vivió hace alrededor de 407.000 años (rango de incertidumbre: 338.000-538.000 años). Otro estimado proporcionó, utilizando suposiciones distintas, una edad mayor: 660.000 años[16].
En aquella época, la especie del género Homo con más presencia en Europa y África era H. heidelbergensis, mientras que en Asia, y en partes de África, existía aun una especie más antigua, H. erectus. Cráneos de estas especies se muestran en la figura. Se puede ver cómo se parecen un poco al neandertal, al tener arcos supraciliares marcados, y una frente baja – H. heidelbergensis más que el neandertal, y H. erectus más que H. heidelbergensis, lo que muestra como en el tiempo se han ido reduciendo estos arcos, y creciendo la frente. En África han sido hallados, además, restos de Homo de la misma época que recibieron el nombre de H. rhodesiensis, mientras que en España vivía H. antecessor.
¡Cuantos nombres! Y a veces pocos huesos, y muy poco ADN preservado, si acaso. ¿Cómo saber si un hallazgo de huesos fósiles pertenece a una, u otra especie? ¿Cómo saber cuál especie dio origen a cuál otra? A menudo la situación no está tan clara como desearíamos. La opinión mayoritaria actual es la plasmada en la figura en este post: H. heidelbergensis es el ancestro común de nosotros y los neandertales[17], y el llamado H. rhodesiensis resultó ser básicamente idéntico a H. heidelbergensis. El ancestro de H. heidelbergensis es H. erectus. Pero hay voces disidentes[18], por ejemplo por el hecho que la evolución de los dientes parece apuntar a un ancestro común de nosotros y los neandertales distinto al heidelbergensis[19], y la última palabra no ha sido hablada.
Queda un asunto por aclarar. Si nuestra especie tiene una edad de unos 200.000 años, y nuestro ancestro común con los neandertales vivió hace 400.000 años o antes, ¿qué pasó en el ínterin entre estas dos fechas? Los descendientes del ancestro común, pertenecientes al parecer a la especie H. heidelbergensis, se separaron en por lo menos dos grupos, uno de los cuales se asentó en el norte (cerca del Mediterráneo), y otro en el sur (África oriental). Del grupo norteño se originaron los neandertales, y del grupo del sur, hace 200.000 años, nosotros.
Adán y Eva
Tal como ya vimos arriba, al comparar el material genético de varias personas se puede determinar cuándo vivió su último ancestro común. Sobre la base de una comparación del ADNmt (el ADN de las mitocondrias, las cuales se transmiten a través de la línea femenina, es decir de madre a hijo/hija), de personas de distintas partes del mundo, se encontró que la “Eva mitocondrial”, la ancestra femenina en términos del ADNmt de todos los seres humanos que viven actualmente, vivió hace unos 200.000 años. Esta edad corresponde bien con la edad de nuestra especie y sugiere que esta “Eva” puede haber sido una de los primeros representantes de H. sapiens. Obviamente, ella no era la única mujer H. sapiens de la época, pero era la única cuyo ADNmt fue transmitido hasta nuestros días[20].
En cuanto a “Adán”, el último ancestro común masculino, la situación es parecida. Un estudio reciente del cromosoma Y, el masculino, indicó que éste pudiera haber vivido mucho antes, hace unos 338.000 años, probablemente en África Central[21]. Sin embargo, se detectó que el análisis en que estaba fundamentada la edad de 338.000 años presentaba algunas fallas debido a unas suposiciones erróneas; un nuevo análisis, utilizando otras suposiciones, entre otras acerca de las frecuencias de mutación, arrojó para “Adán” una edad cercana a los 200.000 años[22], en línea con la edad de nuestra especie. Así que queda confirmada la fecha de nacimiento de nuestra especie. (Esta corrección muestra que es peligroso deducir mucho a partir del material genético uniparental, o sea el cromosoma Y en el caso masculino y el ADN mitocondrial en el femenino. Los análisis actuales toman en consideración todo el material genético de la célula, lo que reduce la probabilidad de conclusiones equivocadas[23].)
Notéis: al hablar de “Adán” y “Eva” no quiero dar la impresión que sólo vivían un hombre y una mujer en aquel tiempo. Más bien había una población de miles de personas, de las cuales sin embargo sólo algunas dejaron descendencia al día de hoy.
Hibridación
Hace unos 60.000 años[24], o antes (ver abajo), nuestros antepasados abandonaron África y se expandieron por todo el planeta, donde entraron en contacto con otras especies de Homo, específicamente nuestros primos: los neandertales y el hombre de Denisova. Inevitable ocurrió, más o menos frecuentemente[25], apareamiento entre nuestros antepasados y aquellos primos. Como resultado, la mayoría de personas en el mundo lleva consigo un pequeño porcentaje de material genético resultante de esta hibridación (producción de crías por padres de dos especies diferentes)[26].
En la actualidad, 1-4 % del material genético de los no africanos se deriva de los neandertales, mientras que este porcentaje es prácticamente nulo en el caso de los africanos[27]. Esto indica que la hibridación de H. sapiens y neandertales tuvo lugar fuera de África, cuando nuestros antepasados ya habían abandonado África en su camino hacia otros continentes. Esto es confirmado por el hecho que la hibridación ocurrió entre 37.000 y 86.000 años antes del presente, o sea, probablemente después de la salida de África[28]. También se encontró en Italia una mandíbula, de hace 30.000-40.000 años, con rasgos mixtos entre neandertal y sapiens, y con un ADNmt (ADN mitocondrial) de neandertal, lo que sugiere hibridación: el individuo pudo haber sido de ascendencia mixta, neandertal-sapiens[29]. (Probablemente se encontraron más restos de individuos híbridos, pero que no fueron reconocidos como tal[30].) De tal manera quedaría rechazada otra hipótesis, que dice que no ocurrió hibridación sino que hubo variaciones genéticas geográficas en la especie ancestral de H. sapiens y los neandertales[31].
Los primeros H. sapiens que llegaron a Asia sur-oriental, se mezclaron – aunque de manera limitada – con sus congéneros de Homo ya asentados allí, relacionados con el hombre de Denisova. Debido a esta hibridación, los pueblos de Australia y el Océano Pacífico tienen hasta 6 % de genes provenientes del hombre de Denisova[32].
Esta hibridación de nuestros antepasados trajo ventajas pequeñas pero claras: el material genético que obtuvimos de los neandertales y los Denisova, incluye genes que refuerzan el sistema inmunológico[33].
Inclusive los pueblos que se quedaron en África, no escaparon cierta hibridación. Hasta en algunos grupos Khoisan (que forman parte de los bosquimanos, quienes están más cerca del H. sapiens “original”) se encontró material genético de otras fuentes: por ejemplo, hasta un 5 % de material genético proveniente del oeste de Eurasia. Esto indica una migración de vuelta hacia África, que tuvo lugar hace unos pocos miles de años[34].
Paradoja de edades
No está completamente claro cuándo empezó el éxodo de H. sapiens desde África. En Israel se encontraron restos de H. sapiens de 90.000 años de antigüedad[35]. En el sur de la China se hicieron varios hallazgos que apuntan a la presencia de H. sapiens por esos lares hace 100.000 a 110.000 años[36]. Esto es significativamente más viejo que la edad de 60.000 años para el éxodo de África basada en los estudios genéticos. O sea, si H. sapiens salió de África sólo hace 60.000 años, ¿cómo es posible que se hayan encontrado restos de lo que parece ser individuos de H. sapiens más viejos fuera de África?
Posibilemente ha sido resuelta esta paradoja: la tasa de mutación del ADN humano pudiera ser más lenta, cerca de la mitad, de lo que se pensaba anteriormente. Por lo tanto, si se estimó a partir de la tasa de mutación previa, una edad de 60.000 años para que el éxodo desde África, este éxodo tal vez ocurrió en realidad hace unos 120.000 años. Si la tasa de mutación más lenta es la correcta, no hay contradicción entre los estudios arqueológicos y genéticos[37]. Sin embargo, dataciones revisadas, basadas en información actualizada acerca de la tasa de mutación del ADNmt humano, colocan la salida desde África al poco tiempo después de 74.000 años antes del presente, así que la paradoja aun no estaría resuelta[38]. De esto tendremos que volver a hablar en un post futuro.
Cuestiones de nombres
Cabe destacar que, si hubo hibridación entre nuestros antepasados y los neandertales y el hombre de Denisova, esto implica que, técnicamente, pertenecemos todos a una misma especie, por lo menos si aceptamos la definición biológica de la especie[39], que es la más comúnmente utilizada. Según esta definición, una especie es un grupo de individuos que pueden cruzarse entre sí, pero que están aislados reproductivamente de otros grupos afines. Al aplicar esta definición de manera estricta, deberíamos considerar a nosotros y a nuestros primos como subespecies de una misma especie, H. sapiens. Nosotros seríamos H. sapiens sapiens, y los neandertales H. sapiens neanderthalensis.
Inclusive es posible que haya más subespecies que las pocas que se han identificado hasta ahora: arriba ya encontramos a H. sapiens idaltu, y, por ejemplo, el hallazgo en Israel de dientes de hace 200.000-400.000 años que parecen más afines a los sapiens que a los neandertales[40] hace pensar que tal vez hubo otros grupos de humanos, pertenecientes de nuestra misma especie pero con variaciones locales.
Conclusión
Según nuestro conocimiento actual, tanto nuestra especie, Homo sapiens, como nuestros “primos”, los neandertales, provenimos de una misma especie antecesora, probablemente Homo heidelbergensis. El hombre de Denisova parece tener los mismos orígenes, mientras que los del hombre de Flores no están tan claras, aunque se piensa que puede haber originado del más antiguo Homo erectus.
Homo sapiens apareció en la África subsahariana, hace unos 200.000 años. Los restos fósiles más antiguos de un H. sapiens anatómicamente moderno (con una edad de 195.000 años) fueron encontrados en Etiopía. Estudios genéticos indican que la ancestra y el ancestro común de todos los humanos actuales vivieron hace alrededor de 200.000 años, y que la población actual que parece ser la más antigua, es la de los bosquimanos en el centro-sur de África.
Así que empezamos poco a poco a tener más claridad acerca del origen de nuestra especie, H. sapiens. Es una historia de migraciones e hibridaciones, lejos de estar desenredada completamente. Lo que está claro ahora es que H. sapiens, aunque más exitoso que las especies, ahora desaparecidas, que recorrieron la tierra cuando él apareció, se mezcló con estas otras especies y les debe algunas características clave.
Pero, ¿qué hizo que desaparecieran las demás especies humanas, y que nosotros, con nuestros orígenes tan humildes, triunfamos? De esto hablaremos en los siguientes posts de esta serie.
Nota: este post fue publicado originalmente, en versión ligeramente distinta, en mi blog – ahora cerrado – “Tiempos de Cambio”. La foto en el encabezado del post es de un Homo sapiens idaltu de hace unos 160.000 años, encontrado en Etiopía (autor: David L. Brill, 2001). Fuente:
http://wadsworth.cengage.com/anthropology_d/special_features/ext/latest_dirt/ethiopia.html.
[1] Ver: http://es.wikipedia.org/wiki/Carlos_Linneo.
[2] Hughes, D.A., Cordaux, R. y Stoneking, M., 2004. Humans. Current Biology, 14 (10), R367-R369. http://www.sciencedirect.com/science/article/pii/S0960982204003276.
[3] Stringer, C.B. y Hublin, J.J., 1999. New age estimates for the Swanscombe hominid and their significance for human evolution. Journal of Human Evolution, 37, 873–877. http://www.eva.mpg.de/evolution/staff/hublin/pdf/Stringer%26Hublin1999.pdf.
[4] Finlayson, C. y otros, 2006. Late survival of Neanderthals at the southernmost extreme of Europe. Nature, 443, 850-853. http://www.nature.com/nature/journal/v443/n7113/full/nature05195.html.
[5] Green, R.E. y otros, 2010. A draft sequence of the Neandertal genome. Science, 328, 710-722. http://www.sciencemag.org/content/328/5979/710.full.pdf.
[6] Meyer, M. y otros, 2012. A high-coverage genome sequence from an Archaic Denisovan individual. Science, 338, 222-226. http://www.sciencemag.org/content/338/6104/222.full.pdf.
[7] Meyer, M. y otros, 2013. A mitochondrial genome sequence of a hominin from Sima de los Huesos. Nature, 505, 403-406. http://www.nature.com/nature/journal/v505/n7483/full/nature12788.html.
[8] Morwood, M.J. y otros, 2004. Archaeology and age of a new hominin from Flores in eastern Indonesia. Nature, 431, 1087-1091. http://www.nature.com/nature/journal/v431/n7012/full/nature02956.html. Brown, P., Sutikna, T., Morwood, M.J., Soejono, R.P., Jatmiko, Wayhu Saptomo, E. y Rokus Awe Due, 2004. A new small-bodied hominin from the Late Pleistocene of Flores, Indonesia. Nature, 431, 1055-1061. http://www.nature.com/nature/journal/v431/n7012/full/nature02999.html.
[9] Ver p.ej.: Baab, K.L., McNulty, K.P. y Harvati, K., 2013. Homo floresiensis contextualized: a geometric morphometric comparative analysis of fossil and pathological human samples. PLoS ONE, 8 (7), e69119. http://www.plosone.org/article/info:doi/10.1371/journal.pone.0069119.
[10] Ver p.ej.: Johanson, D., 2001. Origins of modern humans: multiregional or Out of Africa? Actionbioscience.org. http://www.actionbioscience.org/evolution/johanson.html.
[11] Ver p.ej.: Fagundes, N.J.R., Ray, N., Beaumont, M., Neuenschwander, S., Salzano, F.M., Bonatto, S.L. y Escoffier, L., 2007. Statistical evaluation of alternative models of human evolution. Proceedings of the National Academy of Sciences, 104 (45), 17614-17619. http://intl.pnas.org/content/104/45/17614.full.pdf+html.
[12] Manica, A., Amos, W., Balloux, F. & Hanihara, T., 2007. The effect of ancient population bottlenecks on human phenotypic variation. Nature, 448, 346-348. http://www.nature.com/nature/journal/v448/n7151/full/nature05951.html.
[13] Henn, B.M. et al., 2011. Hunter-gatherer genomic diversity suggests a southern African origin for modern humans. Proceedings of the National Academy of Sciences, 108 (13), 5154-5162. http://www.pnas.org/content/108/13/5154.full.pdf+html.
[14] White, T.D., Asfaw, B., DeGusta, D., Gilbert, H., Richards, G.D., Suwa, G. y Howell, F.C., 2003. Pleistocene Homo sapiens from Middle Awash, Ethiopia. Nature, 423, 742-747. http://www.nature.com/nature/journal/v423/n6941/full/nature01669.html. Clark, J.D. y otros, 2003. Stratigraphic, chronological and behavioural contexts of Pleistocene Homo sapiens from Middle Awash, Ethiopia. Nature, 423, 747-752. http://www.nature.com/nature/journal/v423/n6941/full/nature01670.html. Ver también: Stringer, C., 2003. Out of Ethiopia. Nature, 423, 692-695. http://www.radicalanthropologygroup.org/old/class_text_071.pdf. Hopkin 2003. Skulls reveal dawn of mankind. Nature News. http://www.nature.com/news/2003/030611/full/news030609-8.html. UC Berkeley Press Release, 2003. http://www.berkeley.edu/news/media/releases/2003/06/11_idaltu.shtml. UC Berkeley Press backgrounder, 2003. http://www.berkeley.edu/news/media/releases/2003/06/11_bones-background.shtml.
[15] McDougall, I., Brown, F.H. y Fleagle, J.G., 2005. Stratigraphic placement and age of modern humans from Kibish, Ethiopia. Nature, 433, 733-736. http://www.nature.com/nature/journal/v433/n7027/full/nature03258.html. Aubert, M., Pikeb, A.W.G., Stringer, C., Bartsiokas, A., Kinsley, L., Eggins, S., Day, M. y Grün, R., 2012. Confirmation of a late middle Pleistocene age for the Omo Kibish 1 cranium by direct uranium-series dating. Journal of Human Evolution, 63 (5), 704-710. http://www.sciencedirect.com/science/article/pii/S004724841200142X. Ver también: Hopkin, M., 2005. Ethiopia is top choice for cradle of Homo sapiens. Nature News. http://www.nature.com/news/2005/050216/full/news050214-10.html. Hawks, J., 2005. Omo Kibish redating. http://johnhawks.net/weblog/reviews/early_modern/africa/mcdougall_omo_kibish_2005.html. Lewis, R., 2004. Human origins from Afar. The Scientist. http://www.the-scientist.com/?articles.view/articleNo/15562/title/Human-Origins-from-Afar/.
[16] Stringer, C., 2012. The status of Homo heidelbergensis (Schoetensack 1908). Evolutionary Anthropology, 21, 101-107. http://onlinelibrary.wiley.com/doi/10.1002/evan.21311/pdf. Hublin, J.J., 2009. The origin of Neandertals. Proceedings of the National Academy of Sciences, 106 (38), 16022-16027. http://intl.pnas.org/content/106/38/16022.full.pdf+html.
[17] Mounier, A., Condemi, S. y Manzi, G., 2011. The stem species of our species: a place for the archaic human cranium from Ceprano, Italy. PLoS ONE, 6 (4), e18821. www.plosone.org.
[18] Stringer, 2012 y Hublin, 2009. Ver nota 14.
[19] Gómez-Robles, A., Bermúdez de Castro, J.M., Arsuaga, J.-L., Carbonell, E. y Polly, P.D., 2013. No known hominin species matches the expected dental morphology of the last common ancestor of Neanderthals and modern humans. Proceedings of the National Academy of Sciences, 110 (45), 18196-18201. http://intl.pnas.org/content/110/45/18196.abstract.
[20] Cann, R.L., Stoneking, M. & Wilson, A.C., 1987. Mitochondrial DNA and human evolution. Nature, 325, 31-36. http://www.nature.com/nature/journal/v325/n6099/abs/325031a0.html. Ver también: Cyran, K.A. & Kimmel, M., 2010. Alternatives to the Wright-Fisher model: the robustness of mitochondrial Eve dating. Theoretical Population Biology, 78 (3), 165-172. http://www.sciencedirect.com/science/article/pii/S0040580910000493.
[21] Mendez, F.L. et al., 2013. An African American paternal lineage adds an extremely ancient root to the human Y chromosome phylogenetic tree. American Journal of Human Genetics, 92 (3), 454-459. http://www.cell.com/AJHG/retrieve/pii/S0002929713000736.
[22] Elhaik, E., Tatarinova, T.V., Klyosov, A.A. y Graur, D., 2014. The ‘extremely ancient’ chromosome that isn’t: a forensic bioinformatic investigation of Albert Perry’s X-degenerate portion of the Y chromosome. European Journal of Human Genetics, 22 enero 2014. http://www.nature.com/ejhg/journal/vaop/ncurrent/pdf/ejhg2013303a.pdf.
[23] Lalueza-Fox, C., 2013. Agreements and misunderstandings among three scientific fields. Paleogenomics, archaeology, and human palaeontology. Current Anthropology, 54 (S8), S214-S220. http://www.jstor.org/stable/10.1086/673387.
[24] Henn, B.M., Cavalli-Sforza, L.L. y Feldman, M.W., 2012. The great human expansion. Proceedings of the National Academy of Sciences, 109 (44), 17758-17764. http://intl.pnas.org/content/109/44/17758.full.pdf+html.
[25] Holliday, T.W., Gautney, J.R. y Friedl, L., 2014. Right for the wrong reasons. Reflections on modern human origins in the post-Neanderthal genome era. Current Anthropology, 55 (6), 696-724. http://www.jstor.org/discover/10.1086/679068.
[26] Hammer, M.F., 2013. Human hybrids. Scientific American, mayo 2013, 52-57.
[27] Green y otros, 2010. Ver nota 4.
[28] Sankararaman, S., Patterson, N., Li, H., Pääbo, S. y Reich, D., 2012. The date of interbreeding between Neandertals and modern humans. PLoS Genetics, 8 (10), e1002947. http://www.plosgenetics.org/article/info:doi/10.1371/journal.pgen.1002947.
[29] Condemi, S., Mounier, A., Giunti, P., Lari, M., Caramelli, D. y Longo, L., 2013. Possible interbreeding in late Italian Neanderthals? New data from the Mezzena jaw (Monti Lessini, Verona, Italy). PLoS ONE, 8 (3), e59781. http://www.plosone.org/article/info:doi/10.1371/journal.pone.0059781.
[30] Kelaita, M.A. y Cortés-Ortiz, L., 2013. Morphological variation of genetically confirmed Alouatta Pigra x A. palliata hybrids from a natural hybrid zone in Tabasco, Mexico. American Journal of Physical Anthropology, 150 (2), 223-234. http://onlinelibrary.wiley.com/doi/10.1002/ajpa.22196/abstract.
[31] Eriksson, A. y Manica, A., 2012. Effect of ancient population structure on the degree of polymorphism shared between modern human populations and ancient hominins. Proceedings of the National Academy of Sciences, 109 (35), 13956-13960. http://intl.pnas.org/content/109/35/13956.full.pdf+html.
[32] Meyer y otros, 2012. Ver nota 5.
[33] Abi-Rached, L. et al., 2011. The shaping of the human immune systems by multiregional admixture with archaic humans. Science, 334, 89-94. http://www.sciencemag.org/content/334/6052/89.full.pdf.
[34] Pickrell, J.K., Patterson, N., Loh, P.-R., Lipson, M., Berger, B., Stoneking, M., Pakendorf, B. & Reich, D., 2013. Ancient west Eurasian ancestry in southern and eastern Africa. Preprint. http://arxiv.org/pdf/1307.8014v2.pdf. Gurdasani, D. y otros, 2015. The African genome variation project shapes medical genetics in Africa. Nature, 517, 327-332. http://www.nature.com/nature/journal/v517/n7534/pdf/nature13997.pdf. Lalueza Fox, C., 2012. Los nuevos genomas de homininos del pasado. En: Mateos Cachorro, A. y Perote Alejandre, A. (coords.), 2012. Visiones del ser humano. Del pasado al presente. Instituto Tomás Pascual Sanz, Madrid & CENIEH, Burgos; pág. 61-70. http://www.institutotomaspascualsanz.com/descargas/formacion/publi/Libro_Visiones_del_Ser_Humano.pdf.
[35] Trinkaus, E., 1993. Femoral neck-shaft angles of the Qafzeh-Skhul early modern humans, and activity levels among immature Near Eastern Middle Paleolithic hominids. Journal of Human Evolution, 25 (5), 393–416. http://www.sciencedirect.com/science/article/pii/S0047248483710584.
[36] Shen, G., Wang, W., Wang, Q., Zhao, J., Collerson, K., Zhou, C. y Tobias, P., 2002. U-series dating of Liujiang hominid site in Guangxi, southern China. Journal of Human Evolution, 43 (6), 817-829. http://www.sciencedirect.com/science/article/pii/S0047248402906019. Jin, C.Z. y otros, 2009. The Homo sapiens Cave hominin site of Mulan Mountain, Jiangzhou District, Chongzhuo, Guangxi with emphasis on its age. Chinese Science Bulletin, 54, 3848-3856. http://link.springer.com/article/10.1007/s11434-009-0641-1. Liu, W. et al., 2010. Human remains from Zhirendong, South China, and modern human emergence in East Asia. Proceedings of the National Academy of Sciences, 107 (45), 19201-19206. http://www.pnas.org/content/107/45/19201.full.pdf+html.
[37] Scally, A. & Durbin, R., 2012. Revising the human mutation rate: implications for understanding human evolution. Nature Reviews Genetics, 13, 745-753. http://www.nature.com/nrg/journal/v13/n10/full/nrg3295.html.
[38] Rieux, A. y otros, 2014. Improved calibration of the human mitochondrial clock using ancient genomes. Molecular Biology & Evolution, 31 (10), 2780-2792. http://mbe.oxfordjournals.org/content/31/10/2780.full.pdf+html.
[39] Mayr, E., 1942. Systematics and the origin of species from the viewpoint of a zoologist. Columbia University Press. http://books.google.es/books?id=mAIjnLp6r_MC&printsec=frontcover&hl=es&source=gbs_ge_summary_r&cad=0#v=onepage&q&f=false.
[40] Hershkovitz, I., Smith, P., Sarig, R., Quam, R., Rodríguez, L., García, R., Arsuaga, J.L., Barkai, R. y Gopher, A., 2011. Middle Pleistocene dental remains from Qesem Cave (Israel). American Journal of Physical Anthropology, 144 (4), 575-592. http://onlinelibrary.wiley.com/doi/10.1002/ajpa.21446/abstract.
